

Activation of the atrial KACh channel by the betagamma subunits of G proteins or intracellular Na+ ions depends on the presence of phosphatidylinositol phosphates
- PMID: 9448327
- PMCID: PMC18753
- DOI: 10.1073/pnas.95.3.1307
Activation of the atrial KACh channel by the betagamma subunits of G proteins or intracellular Na+ ions depends on the presence of phosphatidylinositol phosphates
Abstract
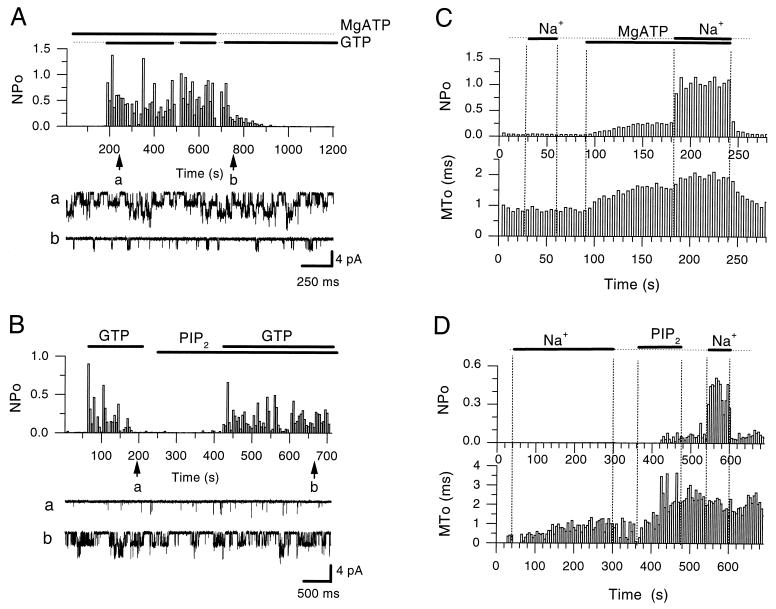
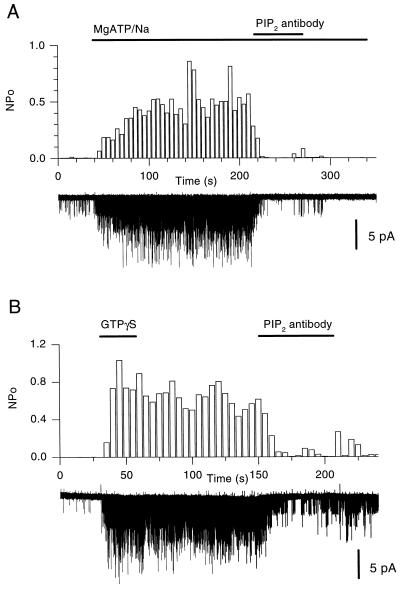
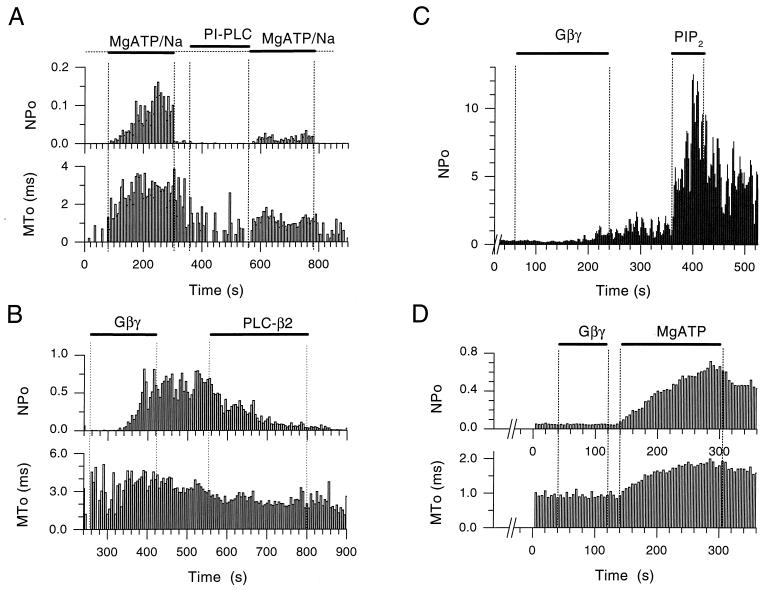
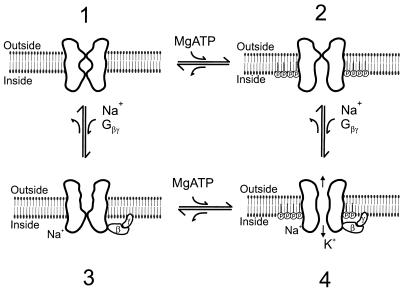
The betagamma subunits of GTP-binding proteins (Gbetagamma) activate the muscarinic K+ channel (KACh) in heart by direct binding to both of its component subunits. KACh channels can also be gated by internal Na+ ions. Both activation mechanisms show dependence on hydrolysis of intracellular ATP. We report that phosphatidylinositol 4,5-bisphosphate (PIP2) mimics the ATP effects and that depletion or block of PIP2 retards the stimulatory effects of Gbetagamma subunits or Na+ ions on channel activity, effects that can be reversed by restoring PIP2. Thus, regulation of KACh channel activity may be crucially dependent on PIP2 and phosphatidylinositol signaling. These striking functional results are in agreement with in vitro biochemical studies on the PIP2 requirement for Gbetagamma stimulation of G protein receptor kinase activity, thus implicating phosphatidylinositol phospholipids as a potential control point for Gbetagamma-mediated signal transduction.
Figures
References
-
- Sui, J.-L., Chan, K., Langan, M.-N., Vivaudou, M. & Logothetis, D. E. in Advances in Second Messenger and Phosphoprotein Research, eds. Armstrong, D. & Rossie, S., in press. - PubMed
-
- Soejima M, Noma A. Pflügers Arch. 1984;400:424–431. - PubMed
-
- Logothetis D E, Kurachi Y, Galper J, Neer E J, Clapham D E. Nature (London) 1987;325:321–326. - PubMed
-
- Krapivinsky G, Gordon E A, Wickman K, Velimirovic B, Krapivinsky L, Clapham D E. Nature (London) 1995;374:135–141. - PubMed
-
- Reuveny E, Slesinger P A, Inglese J, Morales J M, Iniguez-Lluhi J A, Lefkowitz R J, Bourne H R, Jan Y N, Jan L Y. Nature (London) 1994;370:143–146. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
