

Mycoplasma superantigen is a CDR3-dependent ligand for the T cell antigen receptor
- PMID: 9449712
- PMCID: PMC2212128
- DOI: 10.1084/jem.187.3.319
Mycoplasma superantigen is a CDR3-dependent ligand for the T cell antigen receptor
Abstract
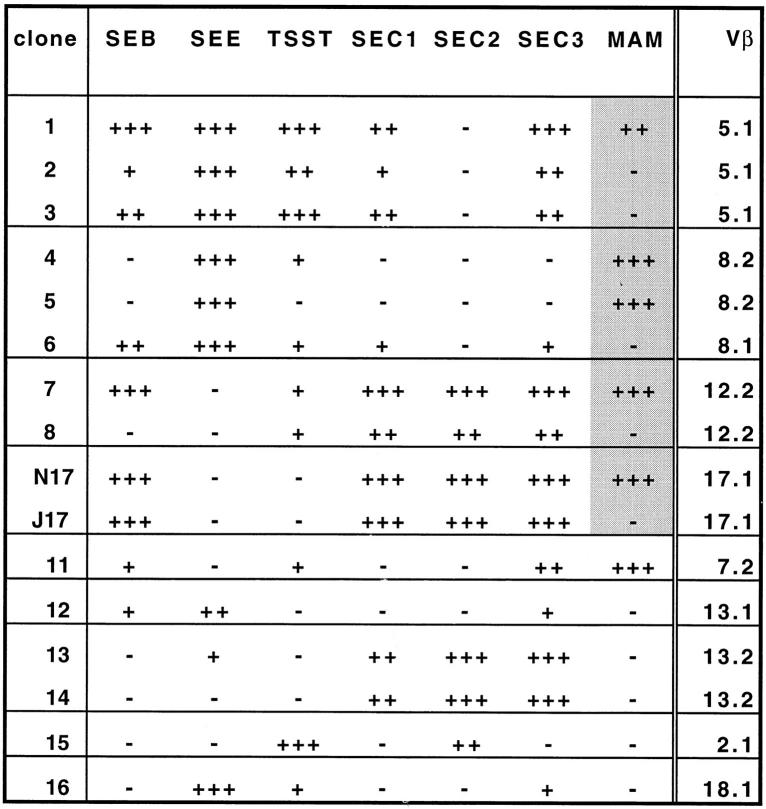
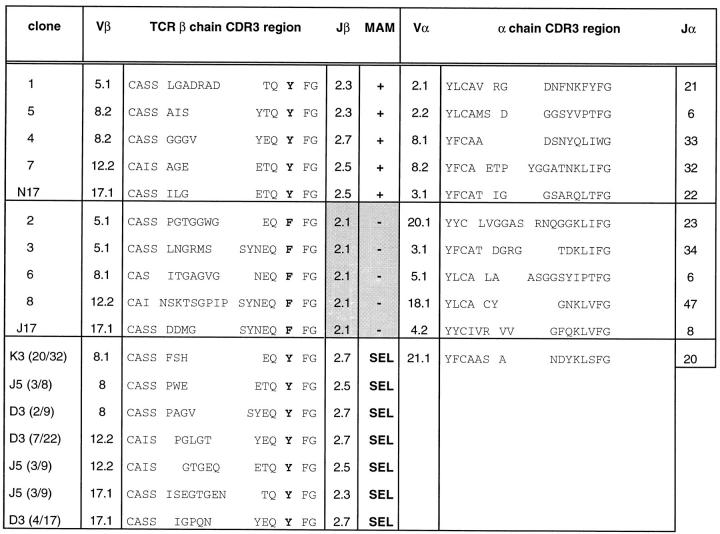
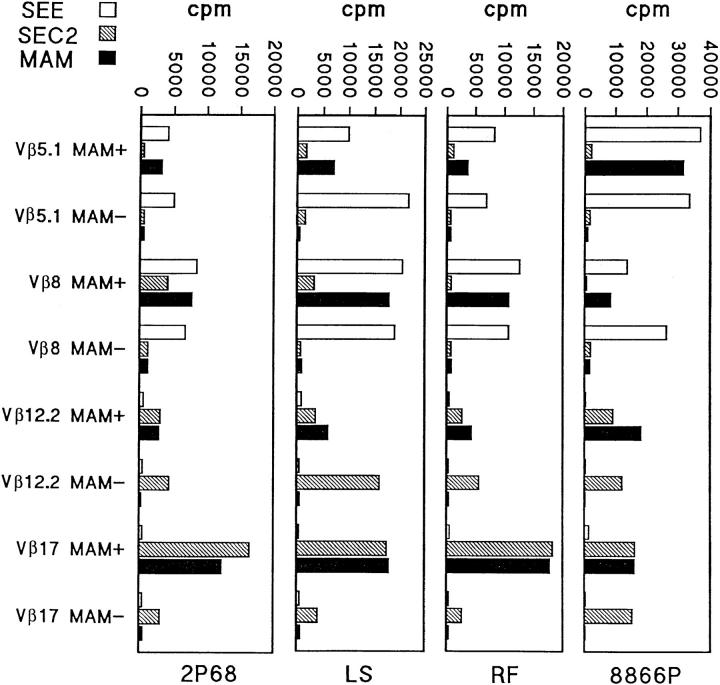
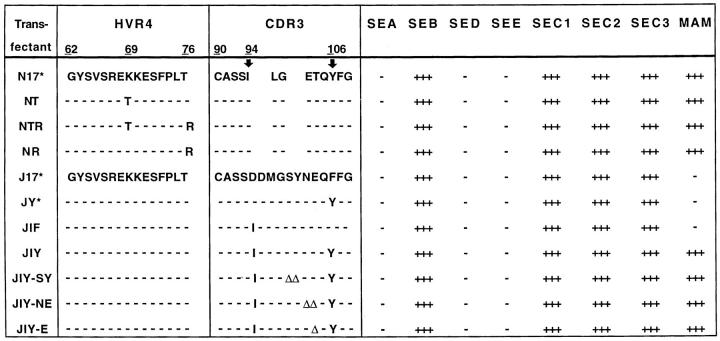
Superantigens are defined as proteins that activate a large number of T cells through interaction with the Vbeta region of the T cell antigen receptor (TCR). Here we demonstrate that the superantigen produced by Mycoplasma arthritidis (MAM), unlike six bacterial superantigens tested, interacts not only with the Vbeta region but also with the CDR3 (third complementarity-determining region) of TCR-beta. Although MAM shares typical features with other superantigens, direct interaction with CDR3-beta is a feature of nominal peptide antigens situated in the antigen groove of major histocompatibility complex (MHC) molecules rather than superantigens. During peptide recognition, Vbeta and Valpha domains of the TCR form contacts with MHC and the complex is stabilized by CDR3-peptide interactions. Similarly, recognition of MAM is Vbeta-dependent and is apparently stabilized by direct contacts with the CDR3-beta region. Thus, MAM represents a new type of ligand for TCR, distinct from both conventional peptide antigens and other known superantigens.
Figures

Similar articles
-
Stimulation of mouse lymphocytes by a mitogen derived from Mycoplasma arthritidis (MAM). VIII. Selective activation of T cells expressing distinct V beta T cell receptors from various strains of mice by the "superantigen" MAM.J Immunol. 1990 Jan 15;144(2):425-31. J Immunol. 1990. PMID: 2136890
-
Human T-cell responses to Mycoplasma arthritidis-derived superantigen.Infect Immun. 1994 Jan;62(1):135-44. doi: 10.1128/iai.62.1.135-144.1994. Infect Immun. 1994. PMID: 8262619 Free PMC article.
-
Interaction of Mycoplasma arthritidis superantigen with human T cells.Clin Infect Dis. 1993 Aug;17 Suppl 1:S170-5. doi: 10.1093/clinids/17.supplement_1.s170. Clin Infect Dis. 1993. PMID: 8399910
-
T-cell receptor structure and TCR complexes.Curr Opin Struct Biol. 1997 Dec;7(6):839-48. doi: 10.1016/s0959-440x(97)80156-x. Curr Opin Struct Biol. 1997. PMID: 9434905 Review.
-
MHC class II-dependent peptide antigen versus superantigen presentation to T cells.Hum Immunol. 1997 May;54(2):194-201. doi: 10.1016/s0198-8859(97)00074-8. Hum Immunol. 1997. PMID: 9297538 Review.
Cited by
-
Attenuation of massive cytokine response to the staphylococcal enterotoxin B superantigen by the innate immunomodulatory protein lactoferrin.Clin Exp Immunol. 2009 Jul;157(1):60-70. doi: 10.1111/j.1365-2249.2009.03963.x. Clin Exp Immunol. 2009. PMID: 19659771 Free PMC article.
-
Association of Mycoplasma arthritidis mitogen with lethal toxicity but not with arthritis in mice.Infect Immun. 2008 Nov;76(11):4989-98. doi: 10.1128/IAI.00667-08. Epub 2008 Sep 8. Infect Immun. 2008. PMID: 18779340 Free PMC article.
-
Mapping the energy of superantigen Staphylococcus enterotoxin C3 recognition of an alpha/beta T cell receptor using alanine scanning mutagenesis.J Exp Med. 2000 Mar 6;191(5):835-46. doi: 10.1084/jem.191.5.835. J Exp Med. 2000. PMID: 10704464 Free PMC article.
-
A general system for targeting MHC class II-antigen complex via a single adaptable loop.Nat Biotechnol. 2024 Dec 13. doi: 10.1038/s41587-024-02466-y. Online ahead of print. Nat Biotechnol. 2024. PMID: 39672953
-
A single point mutation changes the crystallization behavior of Mycoplasma arthritidis-derived mitogen.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006 Mar 1;62(Pt 3):238-41. doi: 10.1107/S1744309106003691. Epub 2006 Feb 10. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006. PMID: 16511311 Free PMC article.
References
-
- Garboczi DN, Ghosh P, Ultz U, Fan QR, Biddison WE, Wiley DC. Structure of the complex between human T-cell receptor, viral peptide and HLA-A2. Nature. 1996;384:134–141. - PubMed
-
- Garcia KC, Degano M, Stanfield RL, Brunmark A, Jackson MR, Peterson PA, Teyton L, Wilson IA. An αβ T cell receptor structure at 2.5 A and its orientation in the TCR–MHC complex. Science. 1996;274:209–219. - PubMed
-
- Schatz DG, Oettinger MA, Schlissel MS. V(D)J recombination: molecular biology and regulation. Annu Rev Immunol. 1992;10:359–383. - PubMed
-
- Janeway CA, Yagi J, Conrad PC, Katz ME, Jones B, Vroegop S, Buxser S. T-cell responses to Mls and to bacterial proteins that mimic its behavior. Immunol Rev. 1989;107:61–88. - PubMed
-
- Marrack P, Kappler J. The staphylococcal enterotoxins and their relatives. Science. 1990;248:705–711. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
