

Accumulation of salicylic acid and 4-hydroxybenzoic acid in phloem fluids of cucumber during systemic acquired resistance is preceded by a transient increase in phenylalanine ammonia-lyase activity in petioles and stems
- PMID: 9449843
- PMCID: PMC35162
- DOI: 10.1104/pp.116.1.231
Accumulation of salicylic acid and 4-hydroxybenzoic acid in phloem fluids of cucumber during systemic acquired resistance is preceded by a transient increase in phenylalanine ammonia-lyase activity in petioles and stems
Abstract
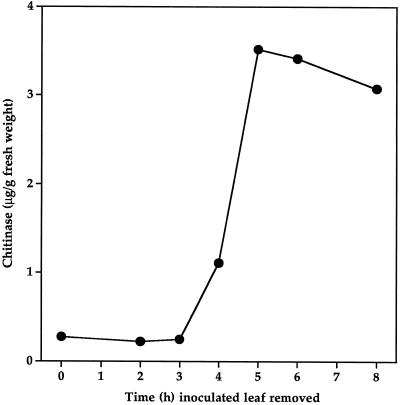
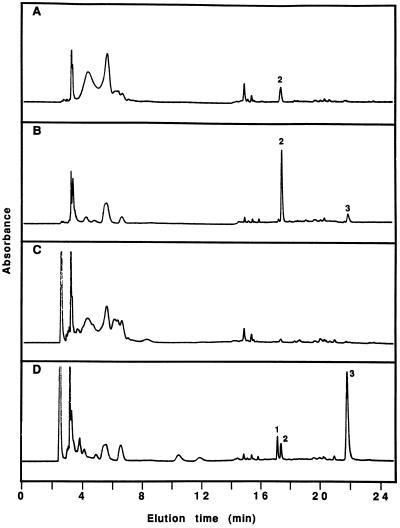
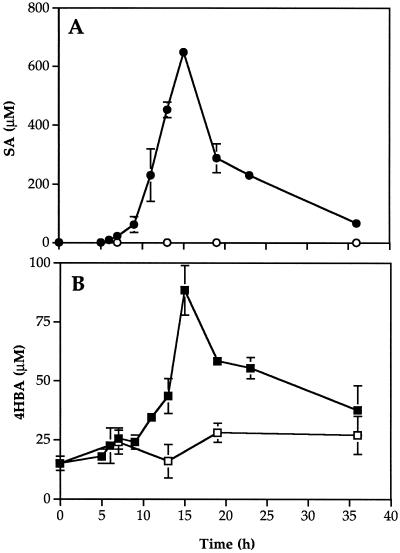
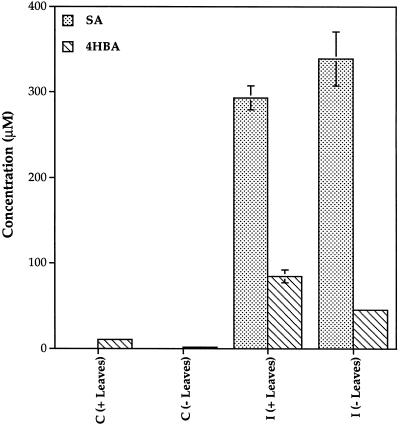
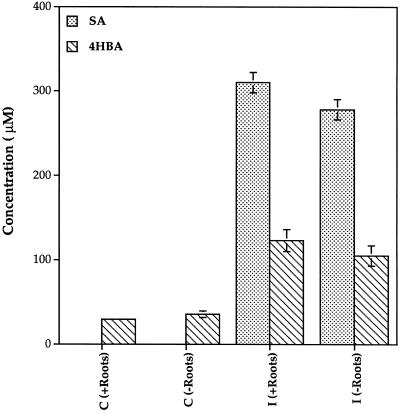
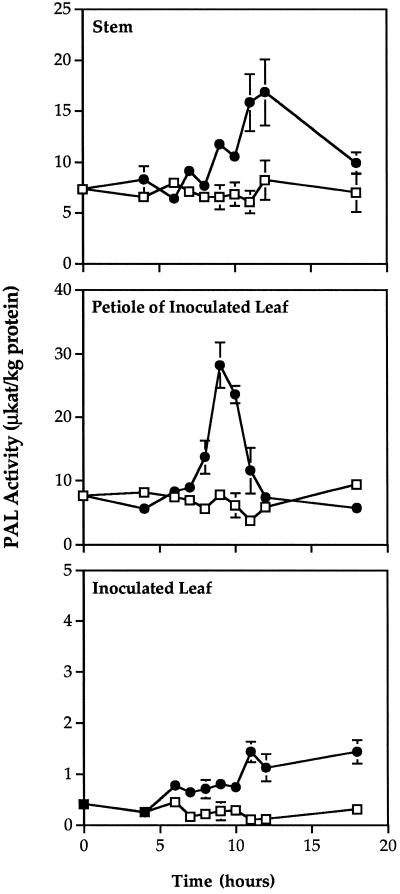
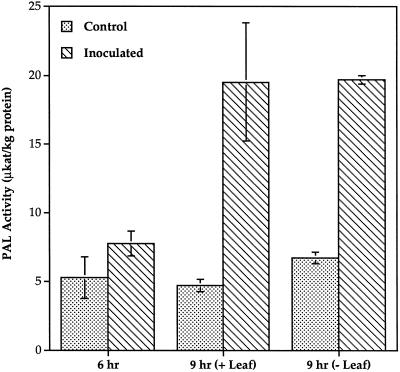
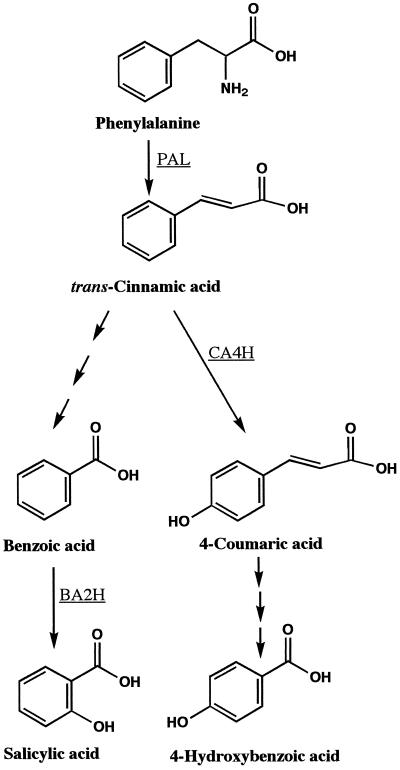
Cucumber (Cucumis sativa) leaves infiltrated with Pseudomonas syringae pv. syringae cells produced a mobile signal for systemic acquired resistance between 3 and 6 h after inoculation. The production of a mobile signal by inoculated leaves was followed by a transient increase in phenylalanine ammonia-lyase (PAL) activity in the petioles of inoculated leaves and in stems above inoculated leaves; with peaks in activity at 9 and 12 h, respectively, after inoculation. In contrast, PAL activity in inoculated leaves continued to rise slowly for at least 18 h. No increases in PAL activity were detected in healthy leaves of inoculated plants. Two benzoic acid derivatives, salicylic acid (SA) and 4-hydroxybenzoic acid (4HBA), began to accumulate in phloem fluids at about the time PAL activity began to increase, reaching maximum concentrations 15 h after inoculation. The accumulation of SA and 4HBA in phloem fluids was unaffected by the removal of all leaves 6 h after inoculation, and seedlings excised from roots prior to inoculation still accumulated high levels of SA and 4HBA. These results suggest that SA and 4HBA are synthesized de novo in stems and petioles in response to a mobile signal from the inoculated leaf.
Figures
References
-
- Boller T, Metraux JP. Extracellular localization of chitinase in cucumber. Physiol Mol Plant Pathol. 1988;33:11–16.
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principles of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Cameron R, Dixon R, Lamb C. Biologically induced systemic acquired resistance in Arabidopsis thaliana. Plant J. 1994;5:715–725.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
