

Ligand-insensitive state of cardiac ATP-sensitive K+ channels. Basis for channel opening
- PMID: 9450949
- PMCID: PMC2222775
- DOI: 10.1085/jgp.111.2.381
Ligand-insensitive state of cardiac ATP-sensitive K+ channels. Basis for channel opening
Abstract
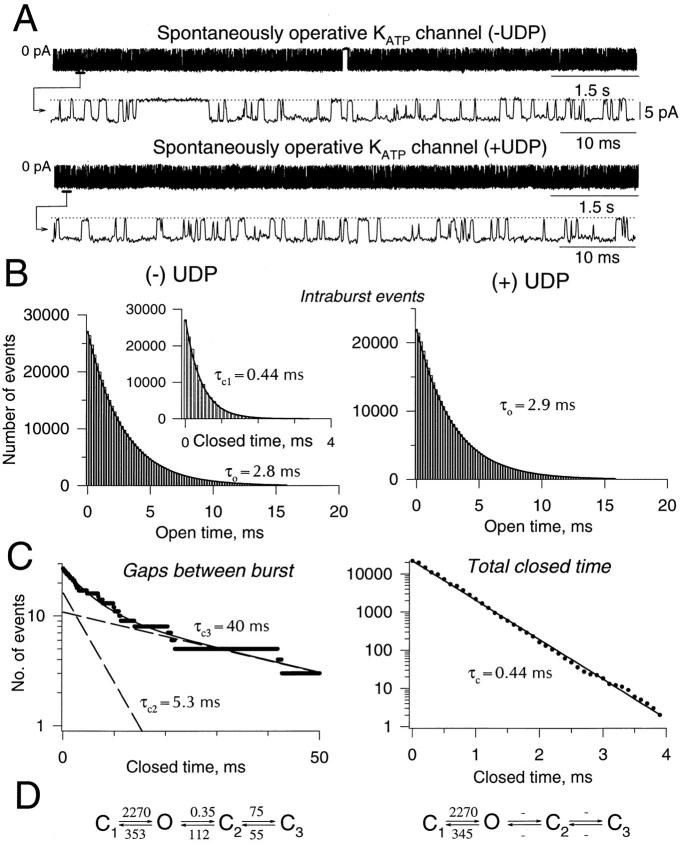
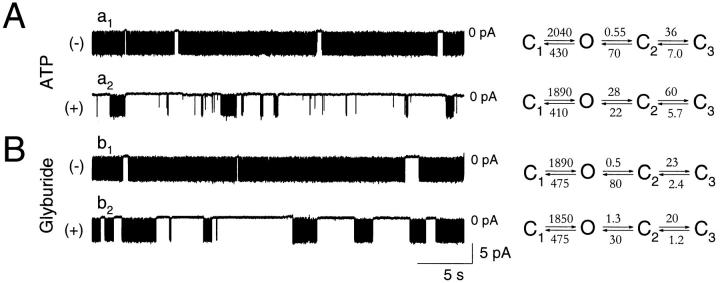
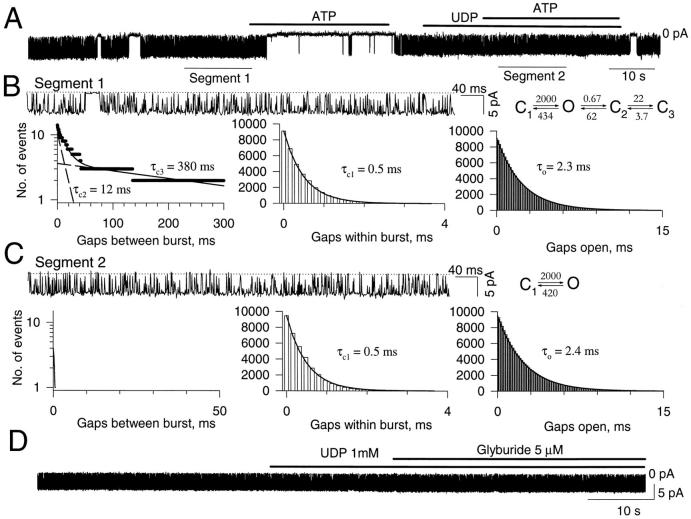
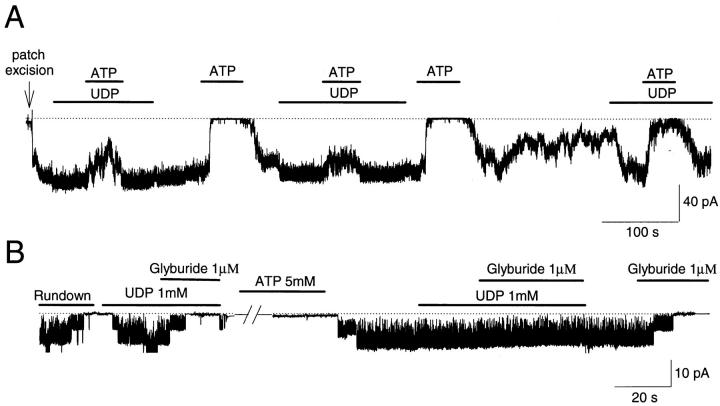
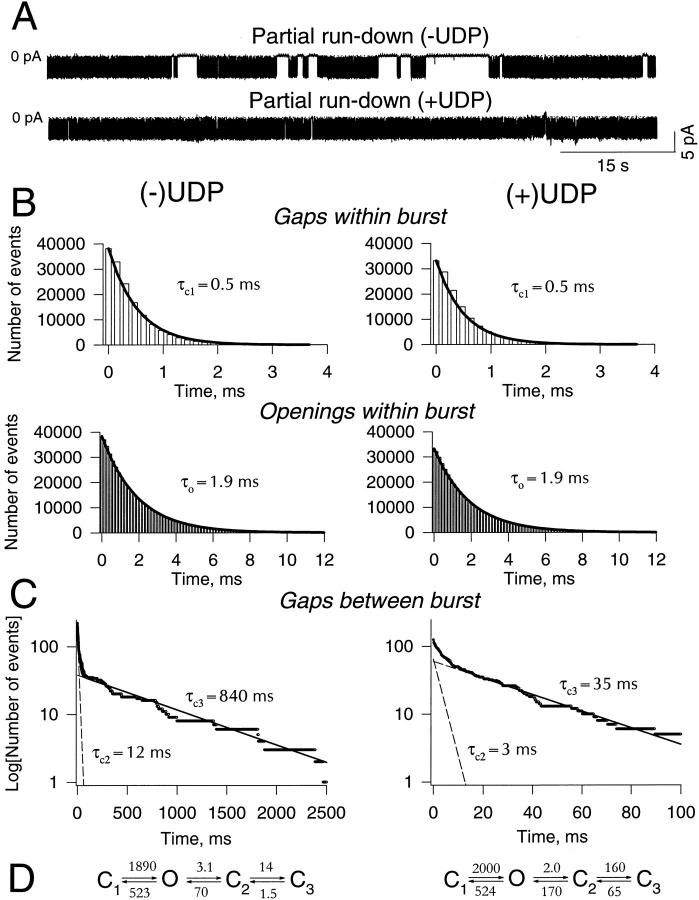
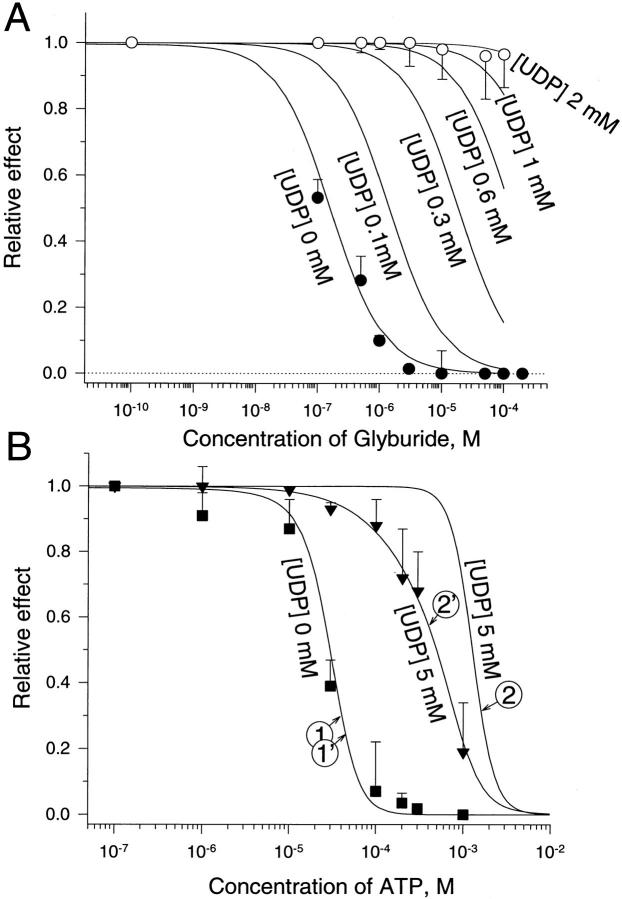
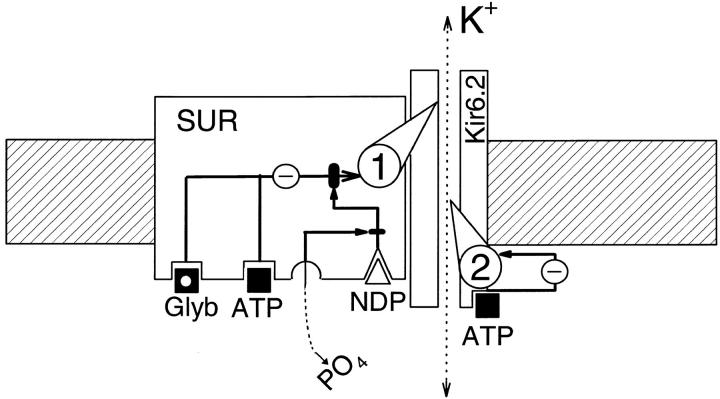
The mechanism by which ATP-sensitive K+ (KATP) channels open in the presence of inhibitory concentrations of ATP remains unknown. Herein, using a four-state kinetic model, we found that the nucleotide diphosphate UDP directed cardiac KATP channels to operate within intraburst transitions. These transitions are not targeted by ATP, nor the structurally unrelated sulfonylurea glyburide, which inhibit channel opening by acting on interburst transitions. Therefore, the channel remained insensitive to ATP and glyburide in the presence of UDP. "Rundown" of channel activity decreased the efficacy with which UDP could direct and maintain the channel to operate within intraburst transitions. Under this condition, the channel was sensitive to inhibition by ATP and glyburide despite the presence of UDP. This behavior of the KATP channel could be accounted for by an allosteric model of ligand-channel interaction. Thus, the response of cardiac KATP channels towards inhibitory ligands is determined by the relative lifetime the channel spends in a ligand-sensitive versus -insensitive state. Interconversion between these two conformational states represents a novel basis for KATP channel opening in the presence of inhibitory concentrations of ATP in a cardiac cell.
Figures
References
-
- Aguilar-Bryan L, Nichols CG, Rajan AS, Parker C, Bryan J. Co-expression of sulfonylurea receptors and KATPchannels in hamster insulinoma tumor (HIT) cells. Evidence for direct association of the receptor with the channel. J Biol Chem. 1992;267:14934–14940. - PubMed
-
- Aguilar-Bryan L, Nichols CG, Wechsler SW, Clement JP, IV, Boyd AE, III, Gonzalez G, Herrera-Sosa H, Nguy K, Bryan J, Nelson DA. Cloning of the beta cell high-affinity sulfonylurea receptor: a regulator of insulin secretion. Science. 1995;268:423–426. - PubMed
-
- Alekseev AE, Gomez LA, Aleksandrova LA, Brady PA, Terzic A. Opening of cardiac sarcolemmal KATPchannels by dinitrophenol separate from metabolic inhibition. J Membr Biol. 1997a;157:203–214. - PubMed
-
- Alekseev AE, Kennedy ME, Navarro B, Terzic A. Burst kinetics of co-expressed Kir6.2/SUR1 clones: comparison of recombinant with native ATP-sensitive K+channel behavior. J Membr Biol. 1997b;159:161–168. - PubMed
-
- Alekseev AE, Jovanovic A, Lopez JR, Terzic A. Adenosine slows the rate of K+-induced membrane depolarization in ventricular cardiomyocytes: possible implication in hyperkalemic cardioplegia. J Mol Cell Cardiol. 1996a;28:1193–1202. - PubMed
