

Evidence for four cytoplasmic dynein heavy chain isoforms in rat testis
- PMID: 9450951
- PMCID: PMC25246
- DOI: 10.1091/mbc.9.2.237
Evidence for four cytoplasmic dynein heavy chain isoforms in rat testis
Abstract
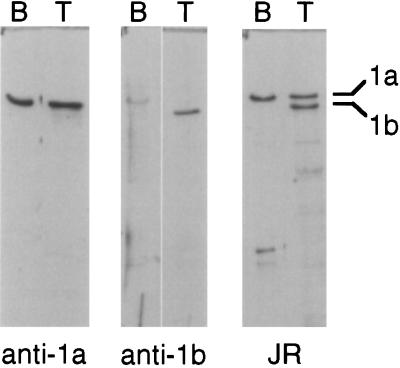
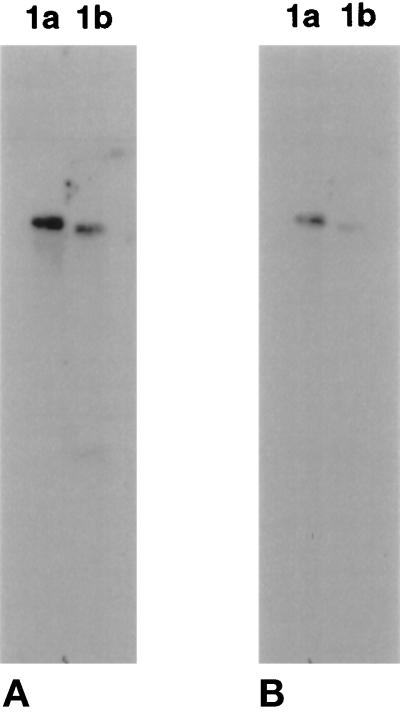
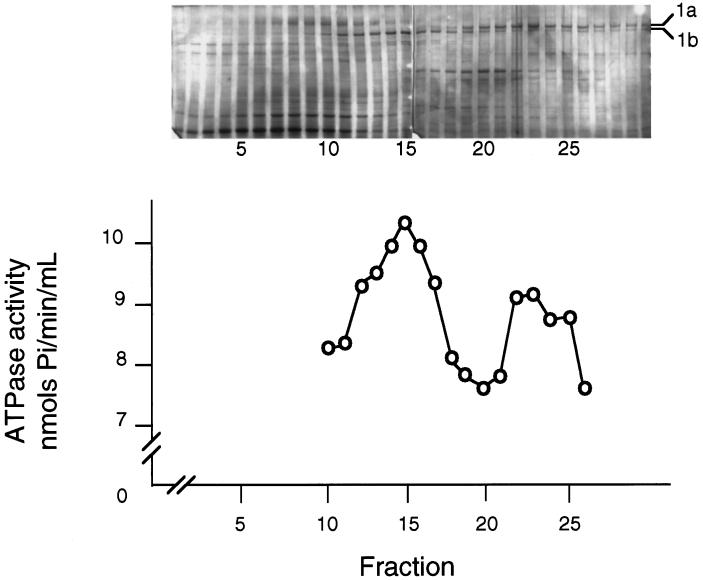
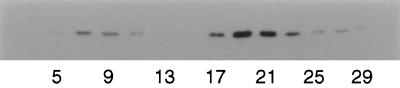
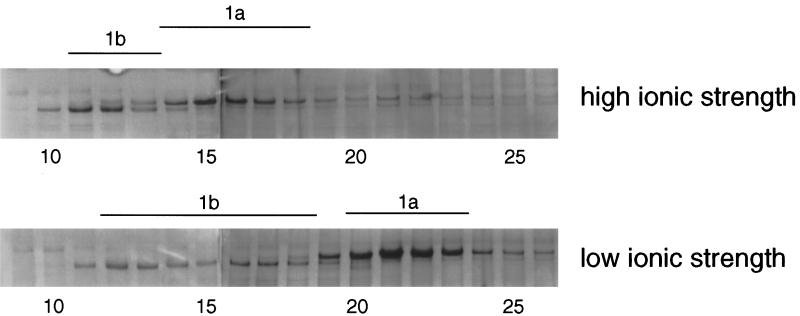
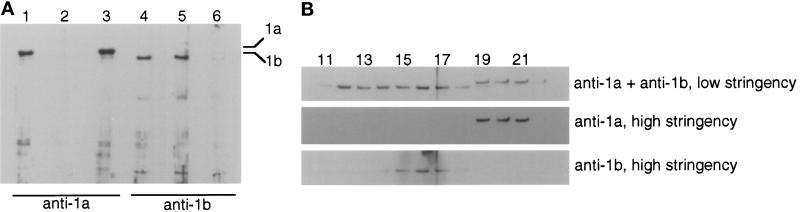
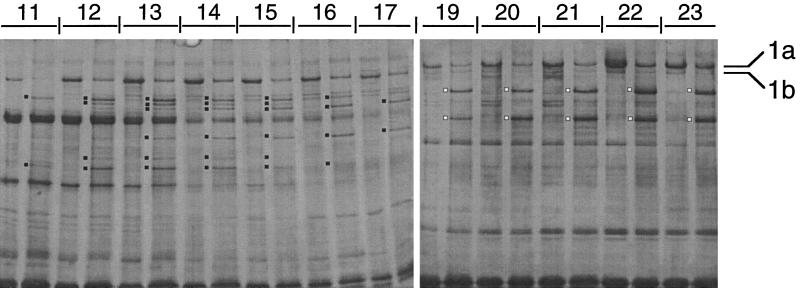
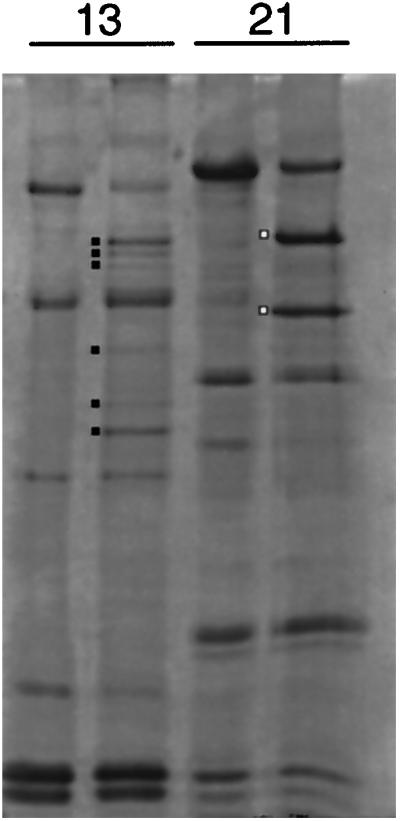
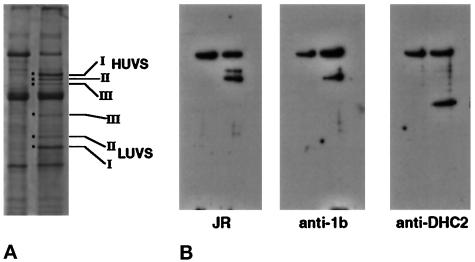
Recent studies have revealed the expression of multiple putative cytoplasmic dynein heavy chain (DHC) genes in several organisms, with each gene encoding a separate protein isoform. This finding is consistent with the hypothesis that different isoforms do different things, as is the case for the axonemal dyneins. Furthermore, the large number of tasks ascribed to cytoplasmic dynein suggests that there may be additional isoforms not yet identified. Two of the mammalian cytoplasmic dynein heavy chains are DHC1a and DHC1b. DHC1a is conventional cytoplasmic dynein and is found in all organisms examined. DHC1b is expressed in organisms that have multiple dyneins, and has been implicated in the intracellular trafficking of molecules in unciliated and ciliated cells. In the present study, we examined the DHC1b protein from rat testis. Testis cytoplasmic dynein contains a large amount of dynein heavy chain reactive with an antibody raised against a peptide sequence of rat DHC1b. The testis anti-DHC1b immunoreactive protein is slightly smaller than testis DHC1a, as assessed by SDS-PAGE. In Northern blots, the DHC1b mRNA is smaller than the DHC1a mRNA. In sucrose gradients made in low ionic strength, DHC1a sedimented at approximately 20S, and the anti-1b immunoreactive heavy chains sedimented in a broad band centered at approximately 14S. The V1-photolysis reaction of individual sucrose gradient fractions revealed three distinct patterns of photolysis, suggesting that there are at least three separate 1b-like heavy chain isoforms in testis. Using a high-stringency Western blotting protocol, the anti-1b antibody and the anti-DHC2 antibody recognized the same heavy chain and specifically bound to one of the three 1b-like heavy chains. We conclude that rat testis contains three 1b-like dynein heavy chains, and one of these is the product of the DHC1b/DHC2 gene previously identified.
Figures
References
-
- Amos LA. Brain dynein crossbridges microtubules into bundles. J Cell Sci. 1989;93:19–28. - PubMed
-
- Asai DJ. Functional and molecular diversity of dynein heavy chains. Semin Cell Dev Biol, 1996;7:311–320.
-
- Asai DJ, Beckwith SM, Kandl KA, Keating HH, Tjandra H, Forney JD. The dynein genes of Paramecium tetraurelia: sequences adjacent to the catalytic P-loop identify cytoplasmic and axonemal heavy chain isoforms. J Cell Sci, 1994;107:839–847. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
