

The synaptic basis of GABAA,slow
- PMID: 9454840
- PMCID: PMC6792721
- DOI: 10.1523/JNEUROSCI.18-04-01305.1998
The synaptic basis of GABAA,slow
Abstract
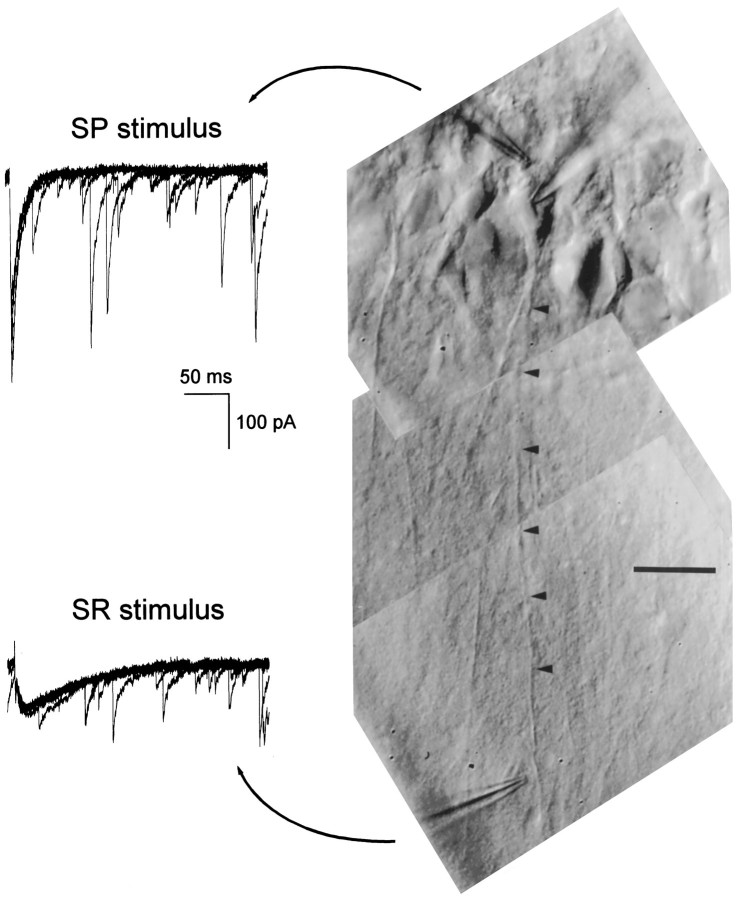
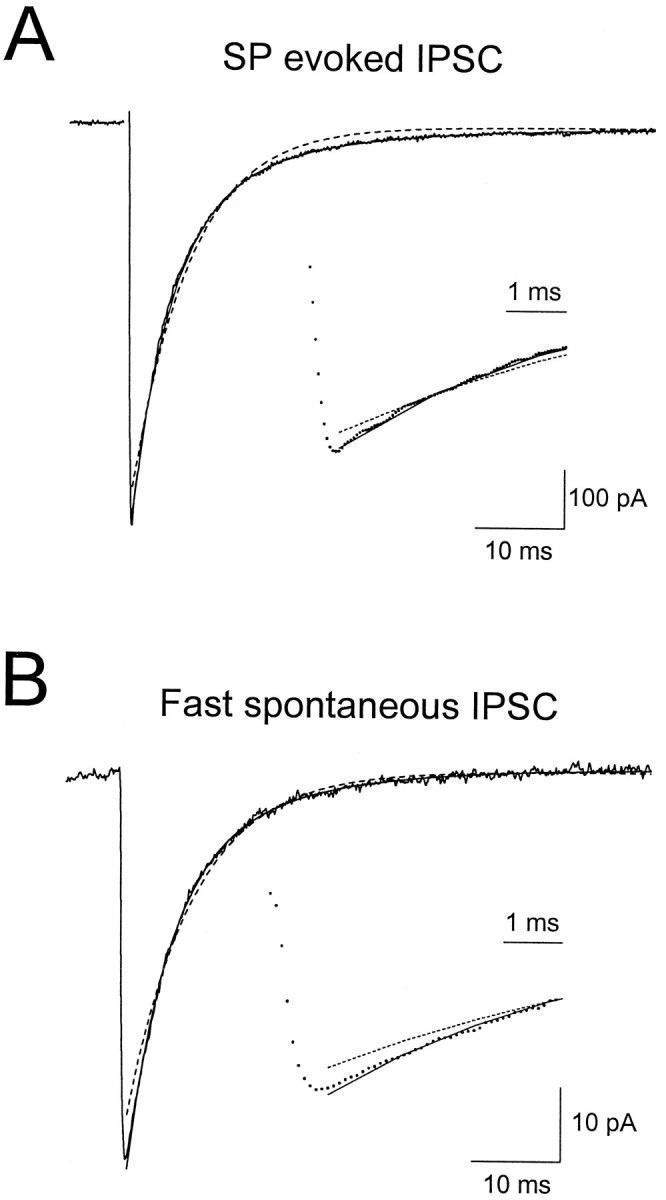
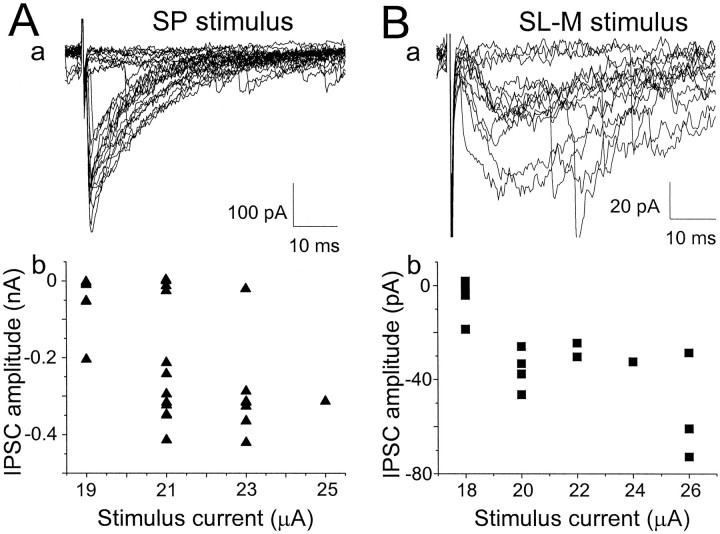
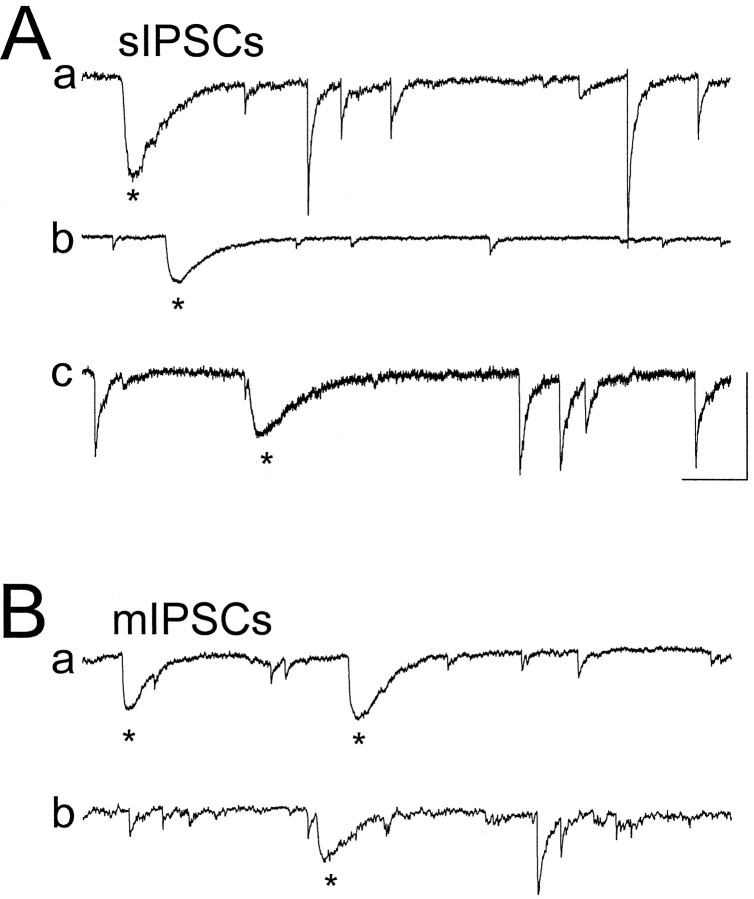
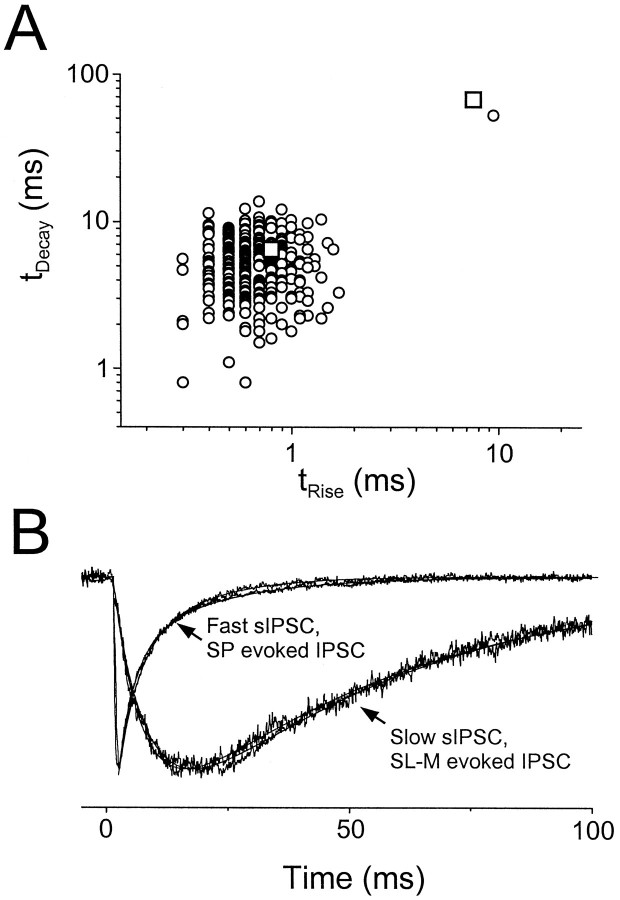
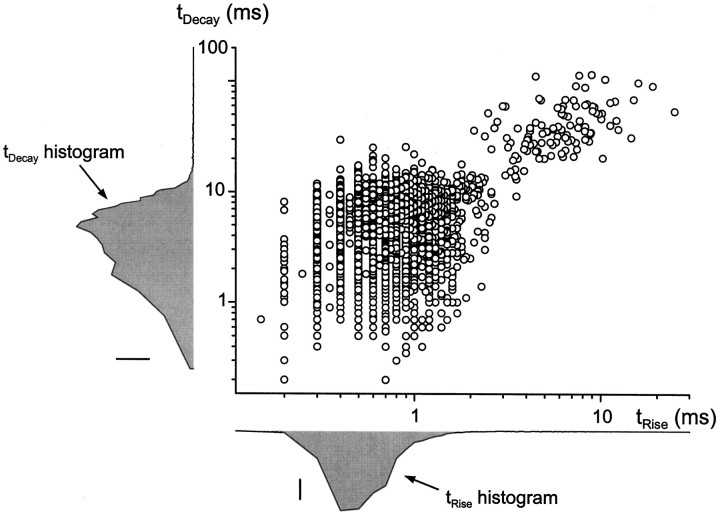
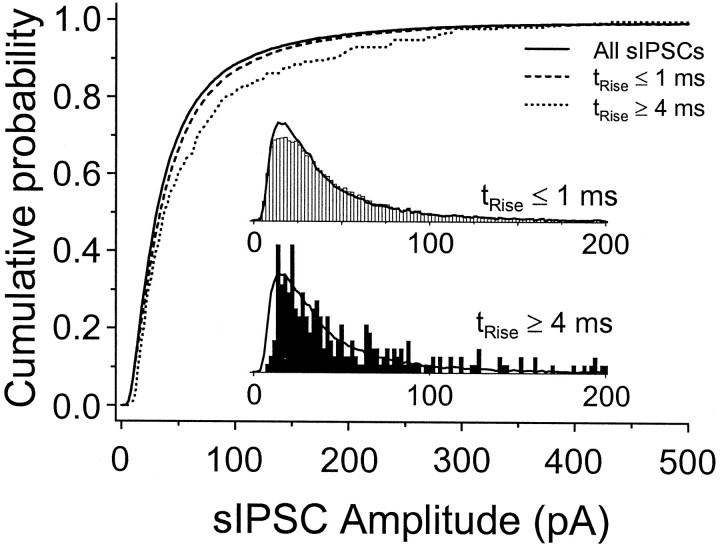
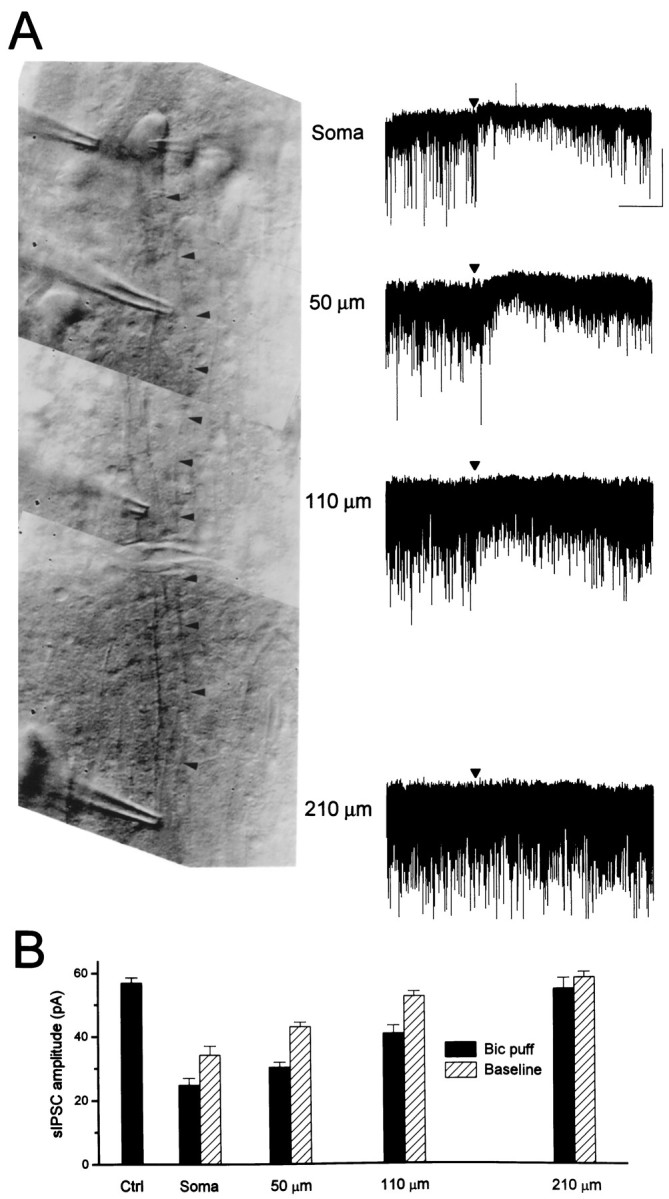
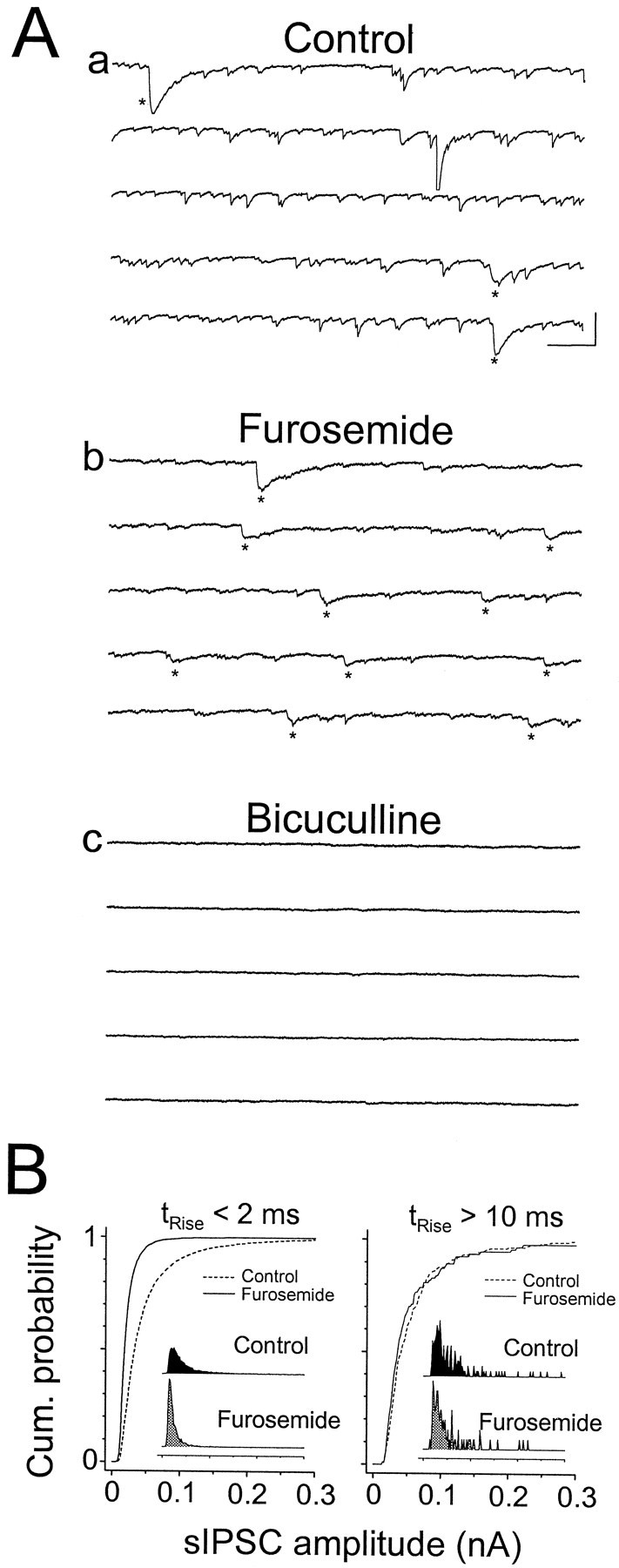
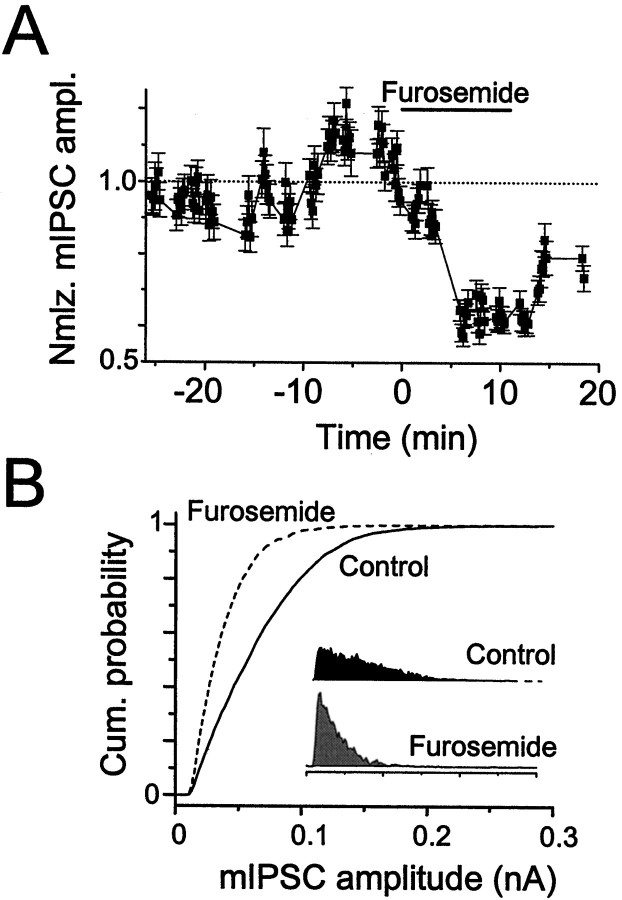
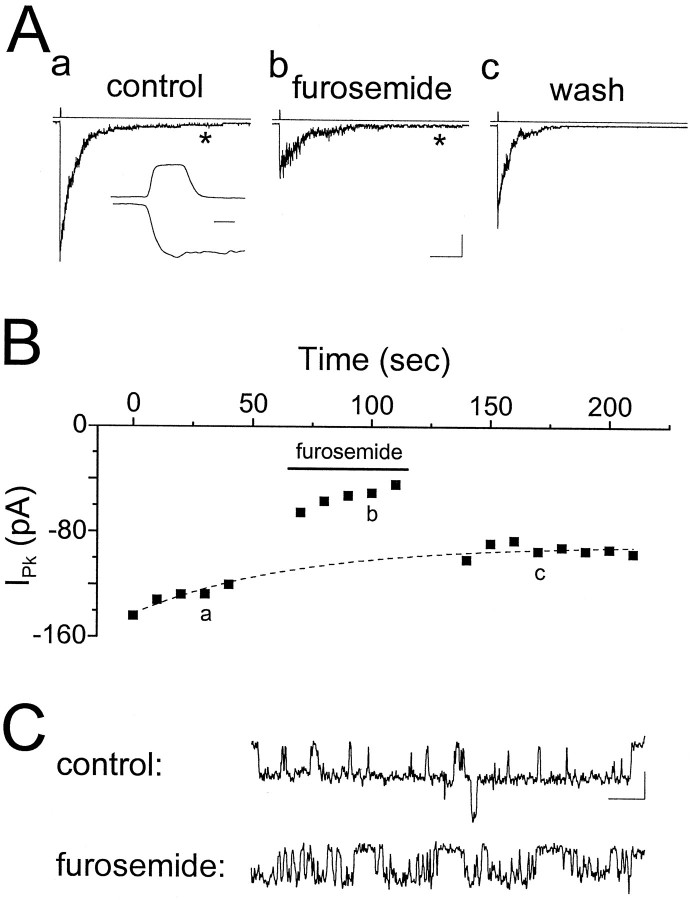
Although two kinetically distinct evoked GABAA responses (GABAA,fast and GABAA,slow) have been observed in CA1 pyramidal neurons, studies of spontaneous IPSCs (sIPSCs) in these neurons have reported only a single population of events that resemble GABAA,fast in their rise and decay kinetics. The absence of slow sIPSCs calls into question the synaptic basis of GABAA,slow. We present evidence here that both evoked responses are synaptic in origin, because two classes of minimally evoked, spontaneous and miniature IPSCs exist that correspond to GABAA,fast and GABAA,slow. Slow sIPSCs occur infrequently, suggesting that the cells underlying these events have a low spontaneous firing rate, unlike the cells giving rise to fast sIPSCs. Like evoked GABAA,fast and GABAA,slow, fast and slow sIPSCs are modulated differentially by furosemide, a subtype-specific GABAA antagonist. Furosemide blocks fast IPSCs by acting directly on the postsynaptic receptors, because it reduces the amplitude of both miniature IPSCs and the responses of excised patches to applied GABA. Thus, in the hippocampus, parallel inhibitory circuits are composed of separate populations of interneurons that contact anatomically segregated and pharmacologically distinct postsynaptic receptors.
Figures
References
-
- Ashwood TJ, Lancaster B, Wheal HV. In vivo and in vitro studies on putative interneurones in the rat hippocampus: possible mediators of feed-forward inhibition. Brain Res. 1984;293:279–291. - PubMed
-
- Banks MI, Pearce RA. Fast and slow spontaneous IPSCs in CA1 pyramidal cells. Soc Neurosci Abstr. 1996;22:810.
-
- Barbour B, Keller BU, Llano I, Marty A. Prolonged presence of glutamate during excitatory synaptic transmission to cerebellar Purkinje cells. Neuron. 1994;12:1331–1343. - PubMed
-
- Buhl EH, Halasy K, Somogyi P. Diverse sources of hippocampal unitary inhibitory postsynaptic potentials and the number of synaptic release sites. Nature. 1994a;368:823–827. - PubMed
-
- Buhl EH, Han ZS, Lorinczi Z, Stezhka VV, Karnup SV, Somogyi P. Physiological properties of anatomically identified axo-axonic cells in the rat hippocampus. J Neurophysiol. 1994b;71:1289–1307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous