

Inhibition of apoptosis in chlamydia-infected cells: blockade of mitochondrial cytochrome c release and caspase activation
- PMID: 9463399
- PMCID: PMC2212145
- DOI: 10.1084/jem.187.4.487
Inhibition of apoptosis in chlamydia-infected cells: blockade of mitochondrial cytochrome c release and caspase activation
Abstract
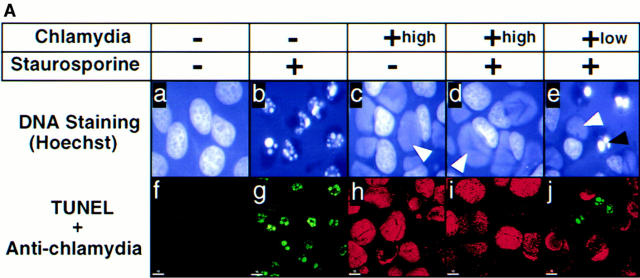
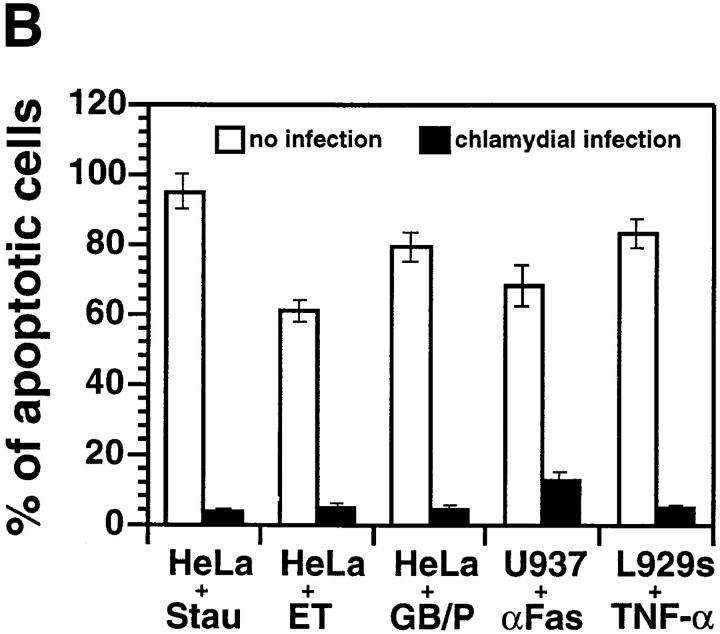
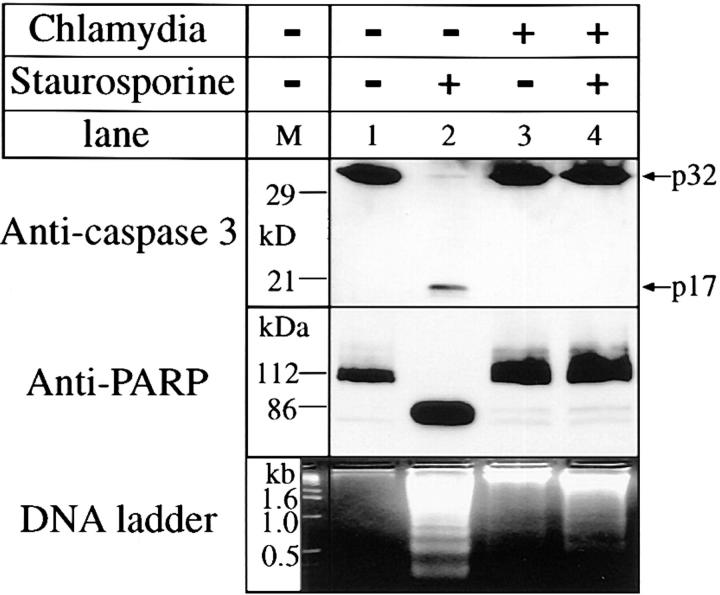
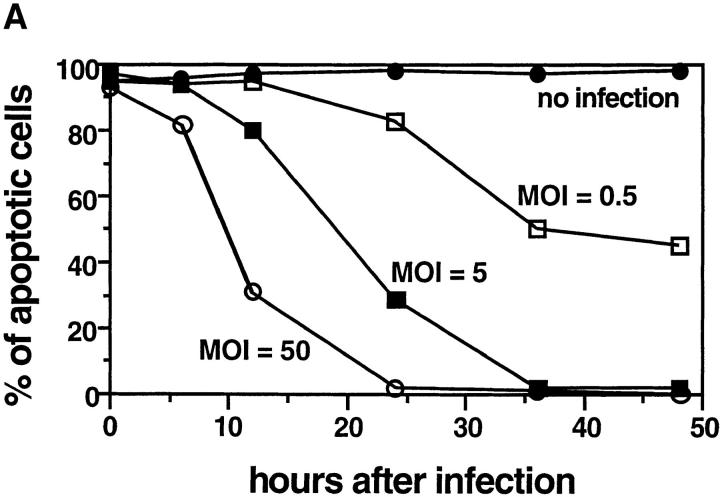
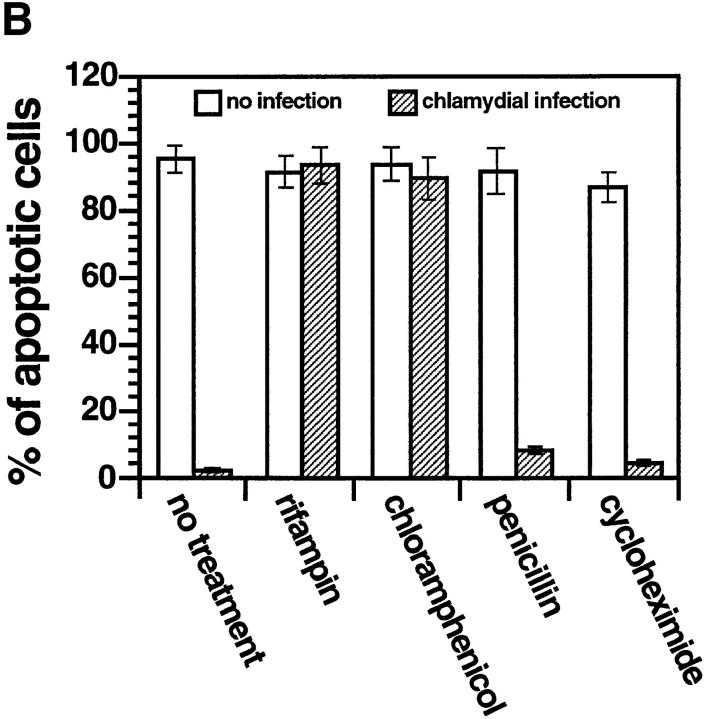
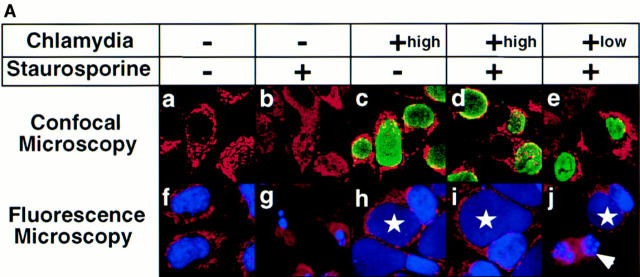
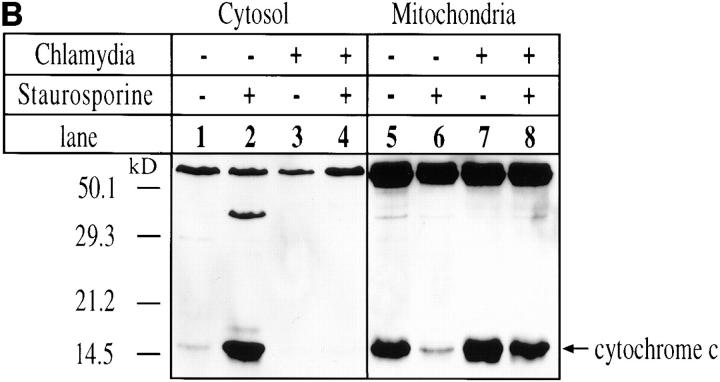
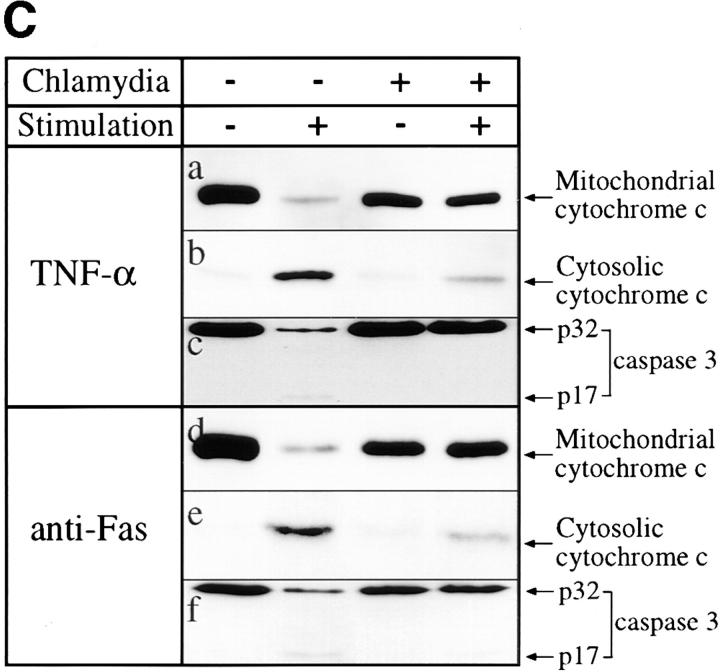
We report that chlamydiae, which are obligate intracellular bacterial pathogens, possess a novel antiapoptotic mechanism. Chlamydia-infected host cells are profoundly resistant to apoptosis induced by a wide spectrum of proapoptotic stimuli including the kinase inhibitor staurosporine, the DNA-damaging agent etoposide, and several immunological apoptosis-inducing molecules such as tumor necrosis factor-alpha, Fas antibody, and granzyme B/perforin. The antiapoptotic activity was dependent on chlamydial but not host protein synthesis. These observations suggest that chlamydia may encode factors that interrupt many different host cell apoptotic pathways. We found that activation of the downstream caspase 3 and cleavage of poly (ADP-ribose) polymerase were inhibited in chlamydia-infected cells. Mitochondrial cytochrome c release into the cytosol induced by proapoptotic factors was also prevented by chlamydial infection. These observations suggest that chlamydial proteins may interrupt diverse apoptotic pathways by blocking mitochondrial cytochrome c release, a central step proposed to convert the upstream private pathways into an effector apoptotic pathway for amplification of downstream caspases. Thus, we have identified a chlamydial antiapoptosis mechanism(s) that will help define chlamydial pathogenesis and may also provide information about the central mechanisms regulating host cell apoptosis.
Figures
References
-
- Shen Y, Shenk TE. Viruses and apoptosis. Curr Opin Genet Dev. 1995;5:105–111. - PubMed
-
- Fratazzi C, Arbeit RD, Carini C, Remold HG. Programmed cell death of Mycobacterium avium serovar4–infected human macrophages prevents the mycobacteria from spreading and induces mycobacterial growth inhibition by freshly added, uninfected macrophages. J Immunol. 1997;158:4320–4327. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
