

Presynaptic nicotinic receptors facilitate monoaminergic transmission
- PMID: 9465015
- PMCID: PMC6792622
- DOI: 10.1523/JNEUROSCI.18-05-01904.1998
Presynaptic nicotinic receptors facilitate monoaminergic transmission
Abstract
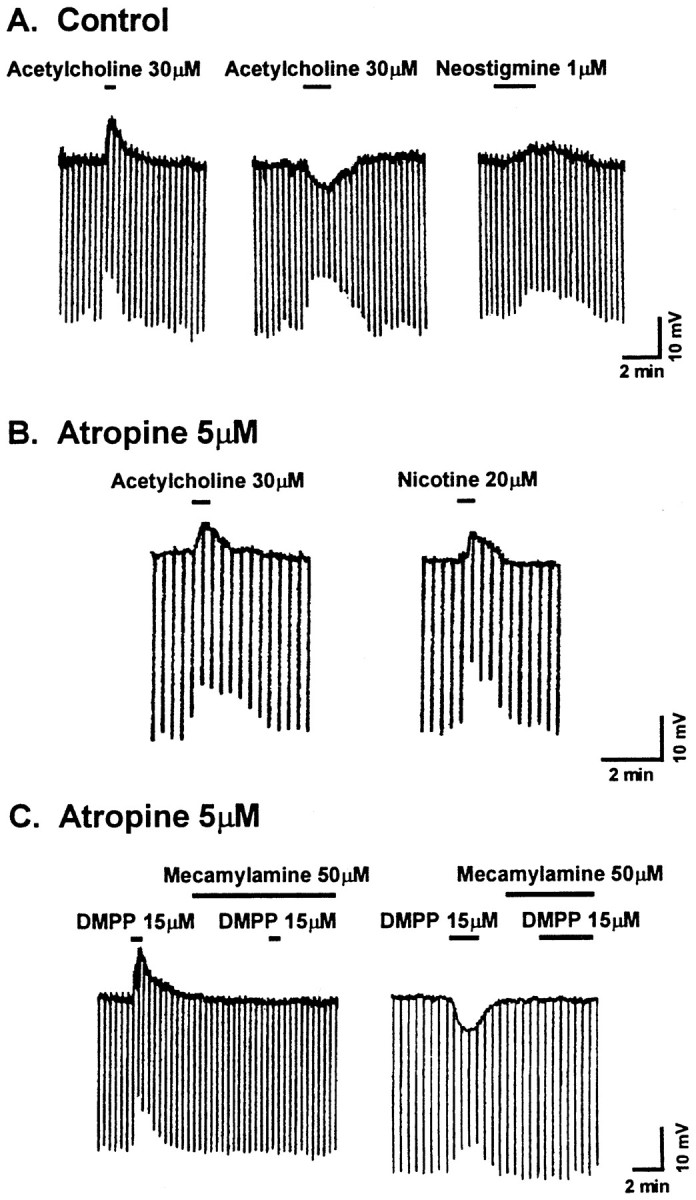
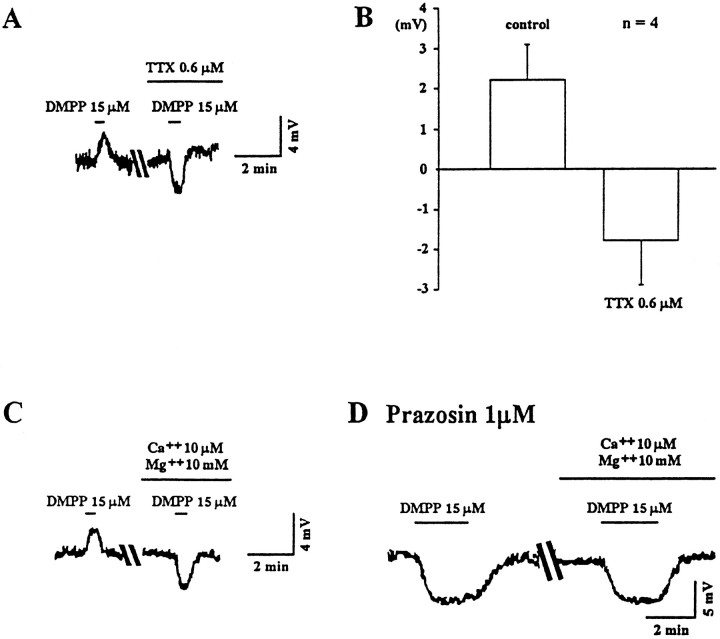
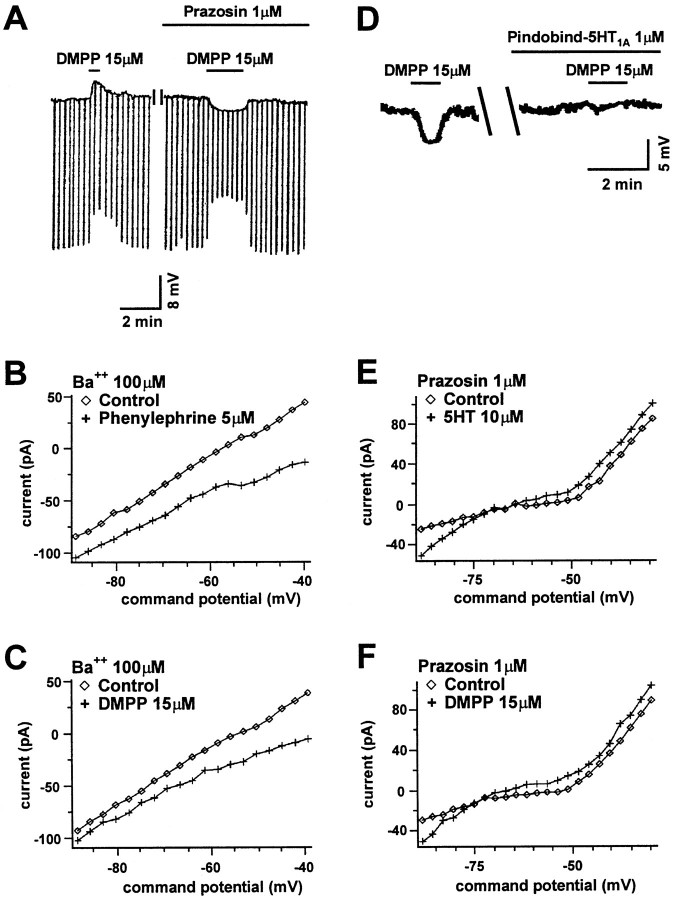
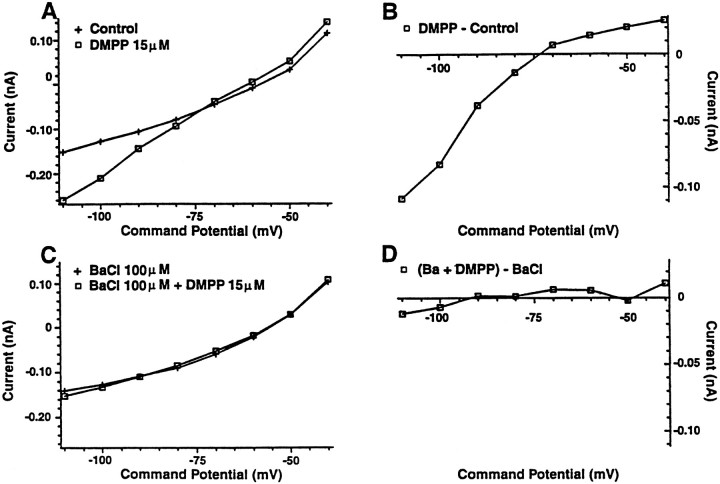
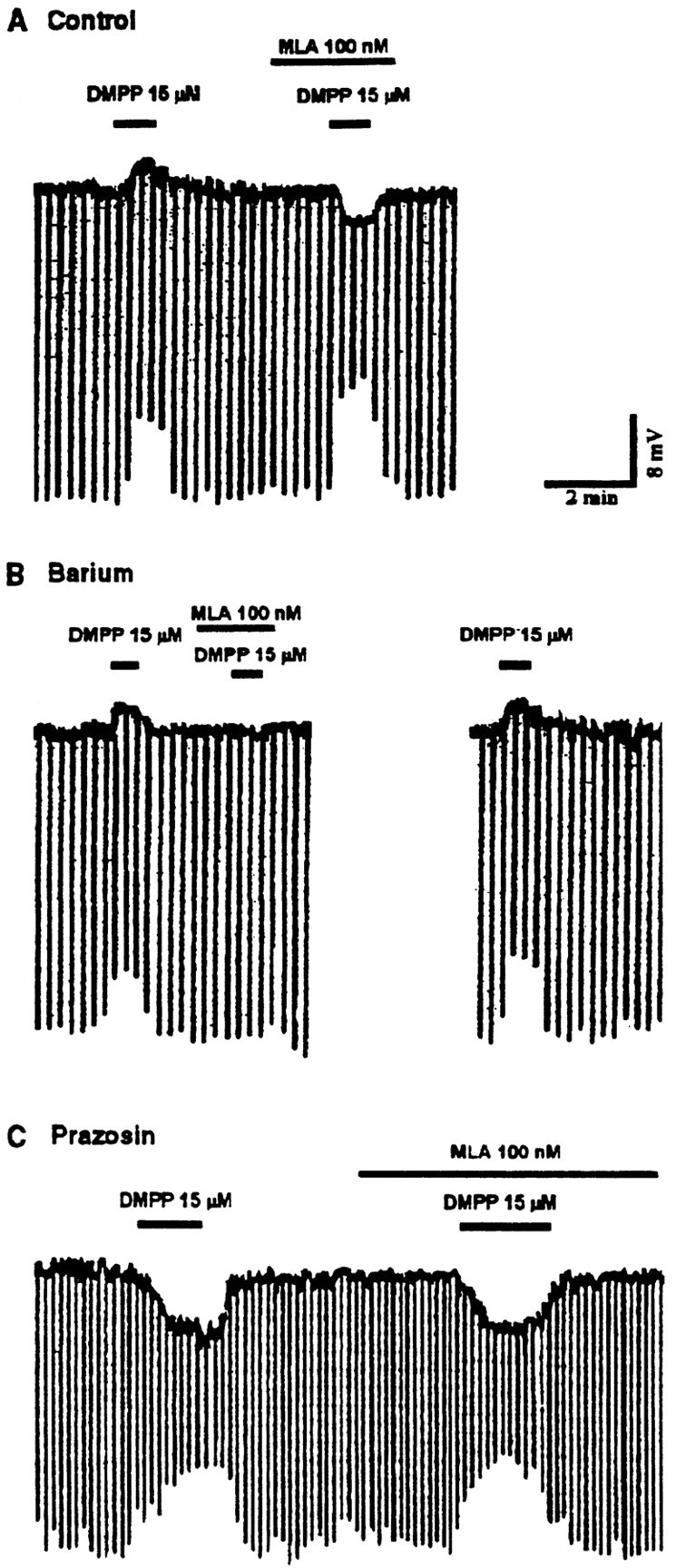
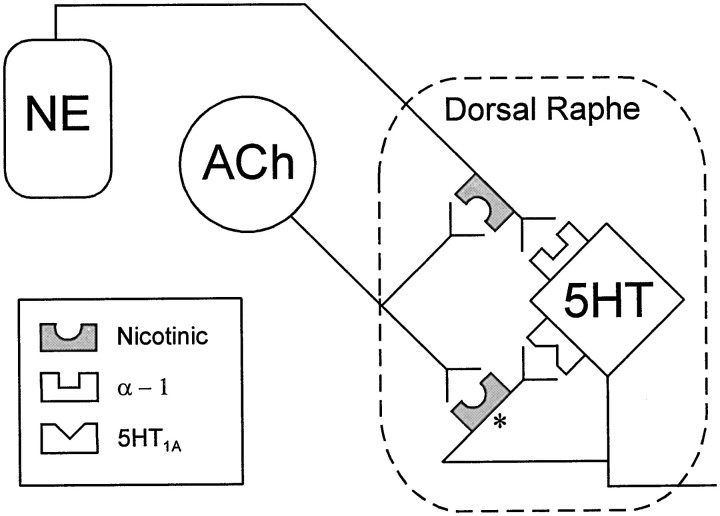
Nicotine is reported to increase arousal and attention and to elevate mood, effects that are most often associated with changes in the function of monoaminergic neuromodulatory systems (Feldman et al., 1997). Recent studies have shown a nicotinic receptor-mediated presynaptic enhancement of fast glutamatergic (McGehee et al., 1995; Gray et al., 1996) and GABAergic (Lena and Changeux, 1997) transmission. However, the mechanism of nicotinic effects on metabotropic-mediated transmission in general, and on monoaminergic transmission in particular, is less well understood. We have examined nicotinic effects on dorsal raphe neurons of rats using whole-cell current and voltage-clamp recording techniques in vitro. In the majority of these neurons, activation of presynaptic nicotinic receptors induced a depolarization mediated by norepinephrine acting on alpha1 receptors. Blockade of this response revealed a hyperpolarization mediated by serotonin acting on 5-HT1A receptors. Because the norepinephrine effect was sensitive to methyllycaconitine (100 nM), it is concluded that nicotinic receptors with an alpha7 subunit can facilitate release of norepinephrine to activate metabotropic receptors. In contrast, methyllycaconitine-insensitive nicotinic receptors can induce 5-HT release in the dorsal raphe nucleus.
Figures

References
-
- Aghajanian GK, Haigler HJ, Bloom FE. Lysergic acid diethylamide and serotonin: direct actions on serotonin-containing neurons in rat brains. Life Sci [I] 1972;11:615–622. - PubMed
-
- Aizenman E, Tang L-H, Reynolds IJ. Effects of nicotinic agonists on the NMDA receptor. Brain Res. 1991;551:355–357. - PubMed
-
- Alkondon M, Albuquerque EX. Diversity of nicotinic acetylcholine receptors in rat hippocampal neurons. I. Pharmacological and functional evidence for distinct structural subtypes. J Pharmacol Exp Ther. 1993;265:1455–1473. - PubMed
-
- Alkondon M, Pereira EF, Wonnacott S, Albuquerque EX. Blockade of nicotinic currents in hippocampal neurons defines methyllycaconitine as a potent and specific receptor antagonist. Mol Pharmacol. 1992;41:802–808. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources