

The hemochromatosis gene product complexes with the transferrin receptor and lowers its affinity for ligand binding
- PMID: 9465039
- PMCID: PMC19050
- DOI: 10.1073/pnas.95.4.1472
The hemochromatosis gene product complexes with the transferrin receptor and lowers its affinity for ligand binding
Abstract
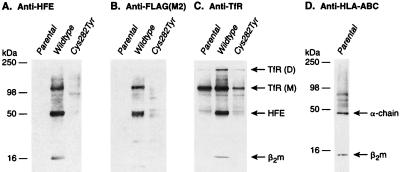
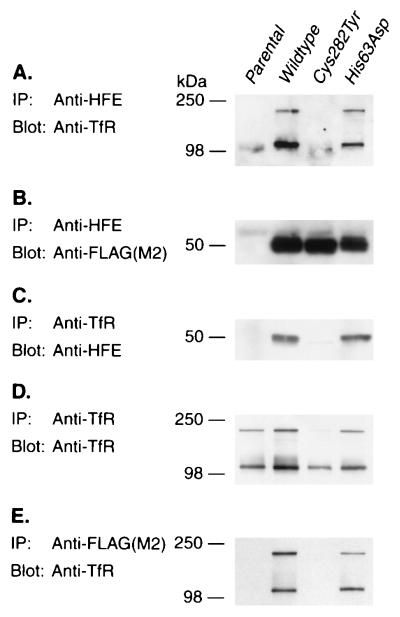
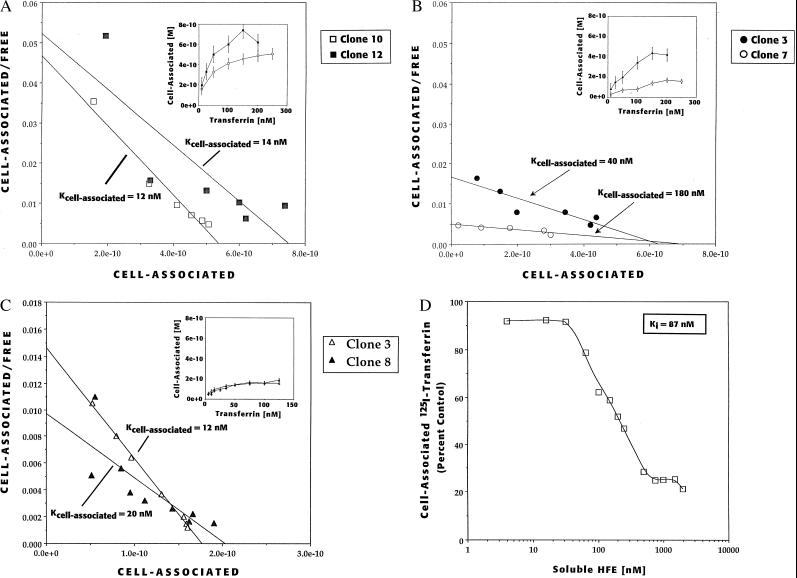
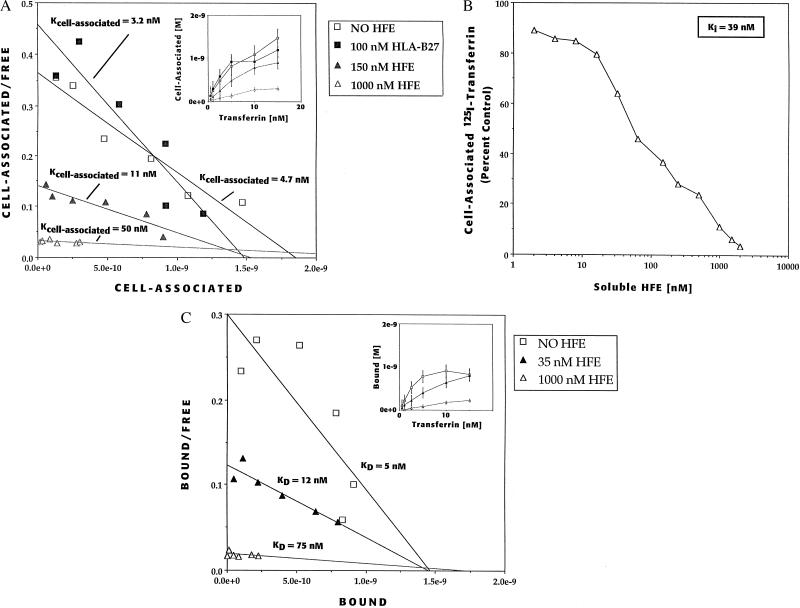
We recently reported the positional cloning of a candidate gene for hereditary hemochromatosis called HFE. The gene product, a member of the major histocompatibility complex class I-like family, was found to have a mutation, Cys-282 --> Tyr (C282Y), in 85% of patient chromosomes. This mutation eliminates the ability of HFE to associate with beta2-microglobulin (beta2m) and prevents cell-surface expression. A second mutation that has no effect on beta2m association, H63D, was found in eight out of nine patients heterozygous for the C282Y mutant. In this report, we demonstrate in cultured 293 cells overexpressing wild-type or mutant HFE proteins that both the wild-type and H63D HFE proteins form stable complexes with the transferrin receptor (TfR). The C282Y mutation nearly completely prevents the association of the mutant HFE protein with the TfR. Studies on cell-associated transferrin at 37 degrees C suggest that the overexpressed wild-type HFE protein decreases the affinity of the TfR for transferrin. The overexpressed H63D protein does not have this effect, providing the first direct evidence for a functional consequence of the H63D mutation. Addition of soluble wild-type HFE/beta2m heterodimers to cultured cells also decreased the apparent affinity of the TfR for its ligand under steady-state conditions, both in 293 cells and in HeLa cells. Furthermore, at 4 degrees C, the added soluble complex of HFE/beta2m inhibited binding of transferrin to HeLa cell TfR in a concentration-dependent manner. Scatchard plots of these data indicate that the added heterodimer substantially reduced the affinity of TfR for transferrin. These results establish a molecular link between HFE and a key protein involved in iron transport, the TfR, and raise the possibility that alterations in this regulatory mechanism may play a role in the pathogenesis of hereditary hemochromatosis.
Figures
References
-
- Dadone M M, Kushner J P, Edwards C Q, Bishop D T, Skolnick M H. Am J Clin Pathol. 1982;78:196–207. - PubMed
-
- Edwards C Q, Griffin L M, Goldgar D, Drummond C, Skolnicx M H, Kushner J P. N Engl J Med. 1988;318:1355–1362. - PubMed
-
- McLaren C E, Gordeuk V R, Looker A C, Hasselblad V, Edward C Q, Griffen L M, Kushner J P, Brittenham G M. Blood. 1995;86:2021–2027. - PubMed
-
- Bothwell T H, Charlton R W, Motulsky A G. In: The Metabolic and Molecular Basis of Inherited Disease. Scriver C R, Beaudet A L, Sly W S, Valle D, editors. New York: McGraw-Hill; 1995. pp. 2237–2269.
-
- Bacon B R, Tavill A S. In: Hepatology. A Textbook of Liver Disease. Zakim D, Boyer T D, editors. Philadelphia: Saunders; 1996. pp. 1439–1472.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
