

Vaccinia NPH-I, a DExH-box ATPase, is the energy coupling factor for mRNA transcription termination
- PMID: 9472022
- PMCID: PMC316528
- DOI: 10.1101/gad.12.4.538
Vaccinia NPH-I, a DExH-box ATPase, is the energy coupling factor for mRNA transcription termination
Abstract
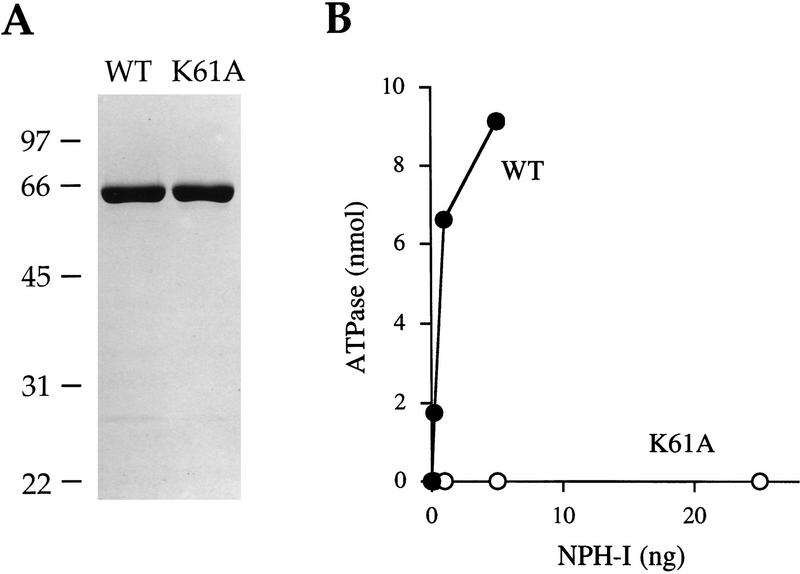
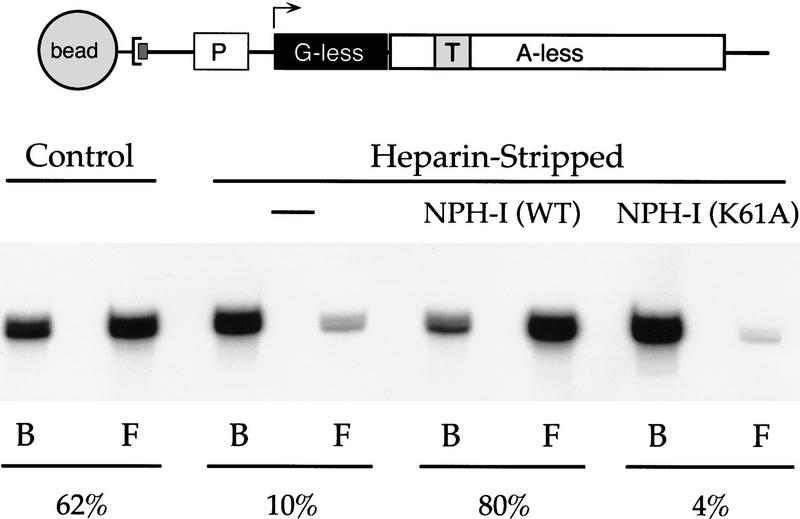
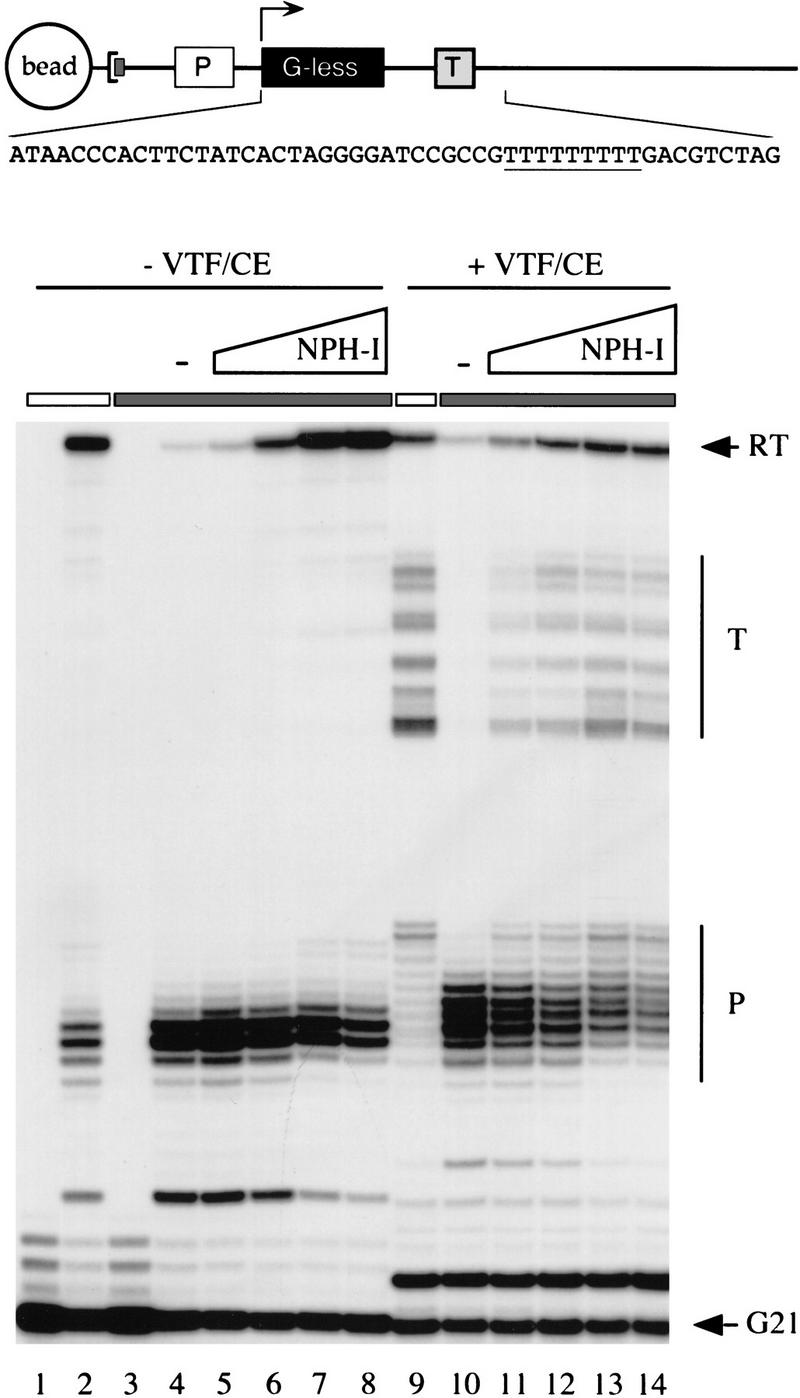
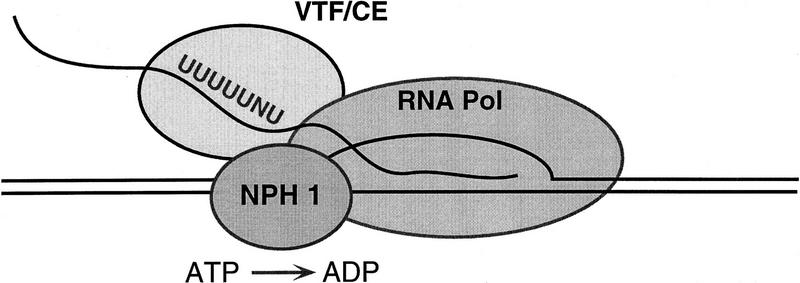
Vaccinia virus RNA polymerase terminates transcription in response to a specific signal UUUUUNU in the nascent RNA. Transduction of this signal to the elongating polymerase requires a trans-acting viral termination factor (VTF/capping enzyme), and is coupled to the hydrolysis of ATP. Recent studies suggest that ATP hydrolysis is catalyzed by a novel termination protein (factor X), which is tightly associated with the elongation complex. Here, we identify factor X as NPH-I (nucleoside triphosphate phosphohydrolase-I), a virus-encoded DNA-dependent ATPase of the DExH-box family. We report that NPH-I serves two roles in transcription (1) it acts in concert with VTF/CE to catalyze release of UUUUUNU-containing nascent RNA from the elongation complex, and (2) it acts by itself as a polymerase elongation factor to facilitate readthrough of intrinsic pause sites. A mutation (K61A) in the GxGKT motif of NPH-I abolishes ATP hydrolysis and eliminates the termination and elongation factor activities. Related DExH proteins may have similar roles at postinitiation steps during cellular mRNA synthesis.
Figures



Similar articles
-
Mutational analysis of vaccinia virus nucleoside triphosphate phosphohydrolase I, a DNA-dependent ATPase of the DExH box family.J Virol. 1999 Feb;73(2):1302-8. doi: 10.1128/JVI.73.2.1302-1308.1999. J Virol. 1999. PMID: 9882335 Free PMC article.
-
Interaction between nucleoside triphosphate phosphohydrolase I and the H4L subunit of the viral RNA polymerase is required for vaccinia virus early gene transcript release.J Biol Chem. 2000 Aug 18;275(33):25798-804. doi: 10.1074/jbc.M002250200. J Biol Chem. 2000. PMID: 10833518
-
Role of forward translocation in nucleoside triphosphate phosphohydrolase I (NPH I)-mediated transcription termination of vaccinia virus early genes.J Biol Chem. 2011 Dec 30;286(52):44764-75. doi: 10.1074/jbc.M111.263822. Epub 2011 Nov 7. J Biol Chem. 2011. PMID: 22069335 Free PMC article.
-
Determinants of vaccinia virus early gene transcription termination.Virology. 2008 Jun 20;376(1):211-24. doi: 10.1016/j.virol.2008.03.011. Epub 2008 Apr 22. Virology. 2008. PMID: 18433825 Free PMC article.
-
Rho-dependent termination and ATPases in transcript termination.Biochim Biophys Acta. 2002 Sep 13;1577(2):251-260. doi: 10.1016/s0167-4781(02)00456-6. Biochim Biophys Acta. 2002. PMID: 12213656 Review.
Cited by
-
Transcriptome view of a killer: African swine fever virus.Biochem Soc Trans. 2020 Aug 28;48(4):1569-1581. doi: 10.1042/BST20191108. Biochem Soc Trans. 2020. PMID: 32725217 Free PMC article. Review.
-
Role of metalloproteases in vaccinia virus epitope processing for transporter associated with antigen processing (TAP)-independent human leukocyte antigen (HLA)-B7 class I antigen presentation.J Biol Chem. 2012 Mar 23;287(13):9990-10000. doi: 10.1074/jbc.M111.314856. Epub 2012 Feb 1. J Biol Chem. 2012. PMID: 22298786 Free PMC article.
-
Transcription apparatus of the yeast virus-like elements: Architecture, function, and evolutionary origin.PLoS Pathog. 2018 Oct 22;14(10):e1007377. doi: 10.1371/journal.ppat.1007377. eCollection 2018 Oct. PLoS Pathog. 2018. PMID: 30346988 Free PMC article.
-
Identification of Vaccinia Virus Replisome and Transcriptome Proteins by Isolation of Proteins on Nascent DNA Coupled with Mass Spectrometry.J Virol. 2017 Sep 12;91(19):e01015-17. doi: 10.1128/JVI.01015-17. Print 2017 Oct 1. J Virol. 2017. PMID: 28747503 Free PMC article.
-
Biochemical and biophysical properties of a putative hub protein expressed by vaccinia virus.J Biol Chem. 2013 Apr 19;288(16):11470-81. doi: 10.1074/jbc.M112.442012. Epub 2013 Mar 8. J Biol Chem. 2013. PMID: 23476017 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources