

Cytoskeletal polarization of T cells is regulated by an immunoreceptor tyrosine-based activation motif-dependent mechanism
- PMID: 9472038
- PMCID: PMC2141749
- DOI: 10.1083/jcb.140.4.861
Cytoskeletal polarization of T cells is regulated by an immunoreceptor tyrosine-based activation motif-dependent mechanism
Abstract
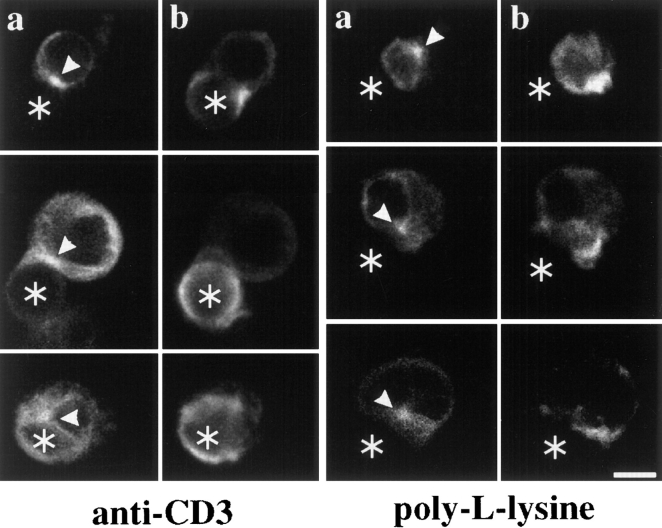
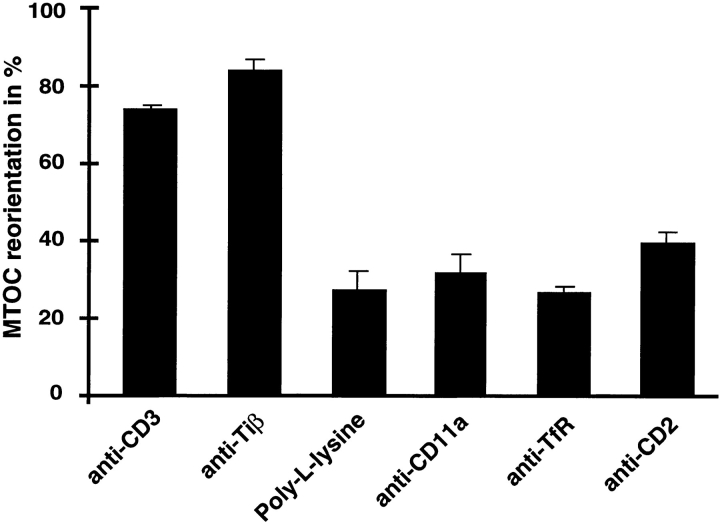
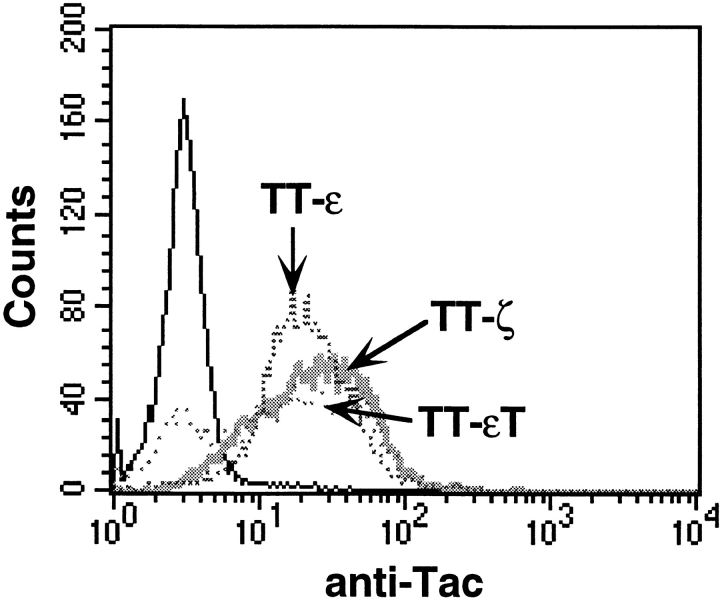
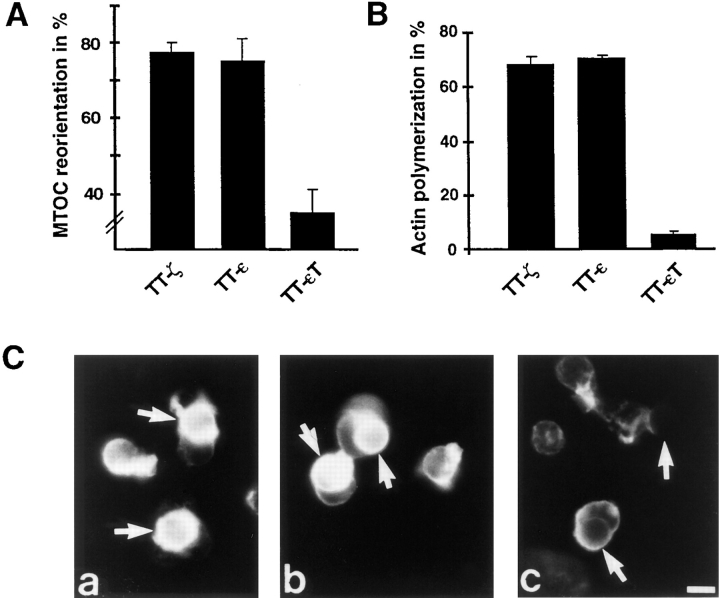
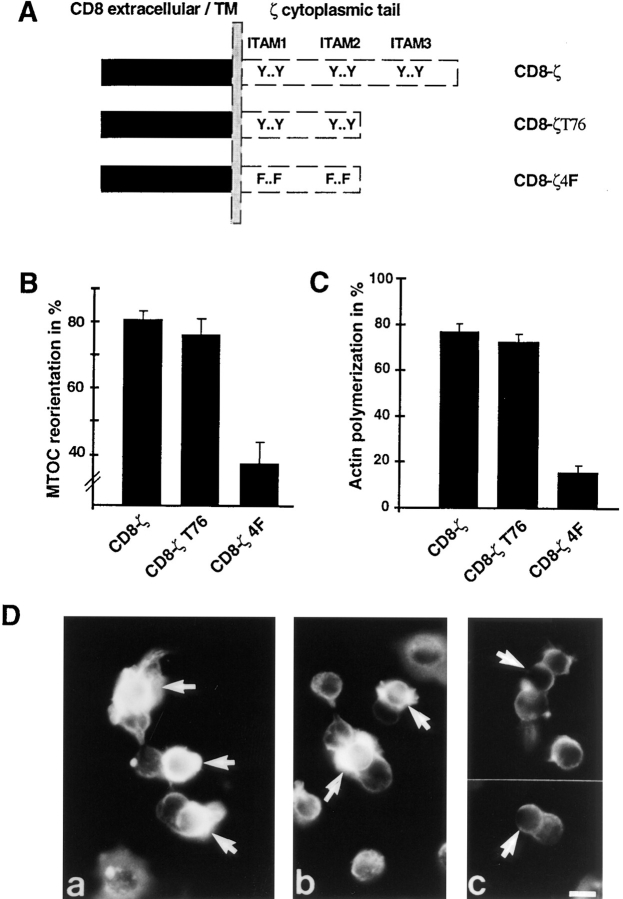
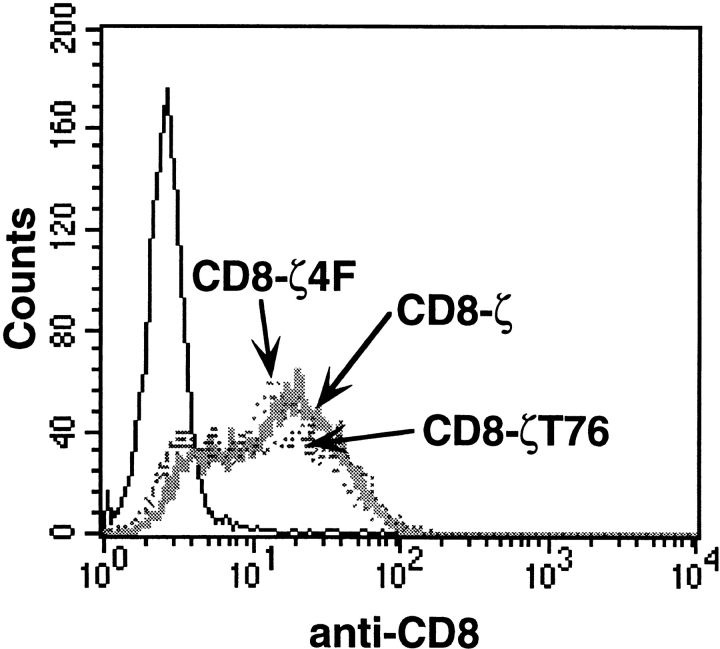
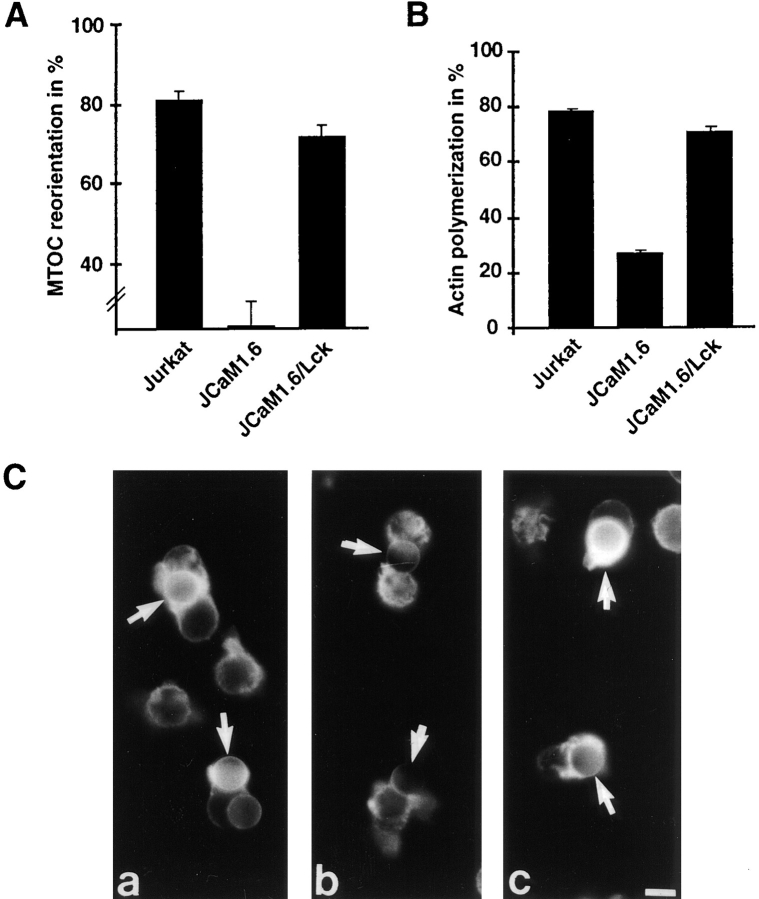
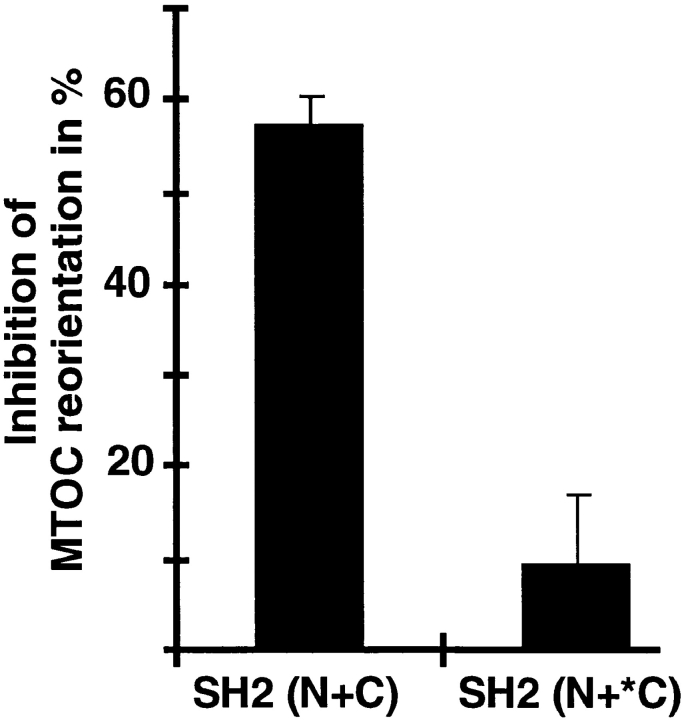
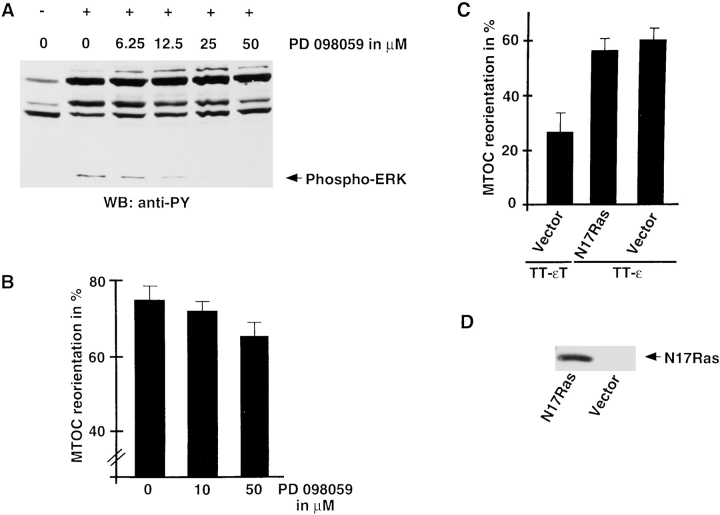
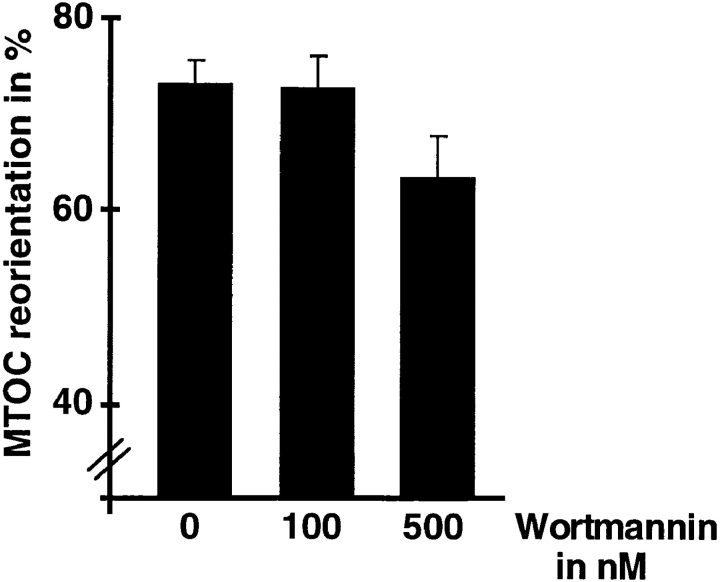
Binding of a T cell to an appropriate antigen-presenting cell (APC) induces the rapid reorientation of the T cell cytoskeleton and secretory apparatus towards the cell-cell contact site in a T cell antigen receptor (TCR) and peptide/major histocompatibility complex-dependent process. Such T cell polarization directs the delivery of cytokines and cytotoxic mediators towards the APC and contributes to the highly selective and specific action of effector T cells. To study the signaling pathways that regulate cytoskeletal rearrangements in T lymphocytes, we set up a conjugate formation assay using Jurkat T cells as effectors and cell-sized latex beads coated with various antibodies as artificial APCs. Here, we report that beads coated with antibodies specific for the TCR-CD3 complex were sufficient to induce T cell polarization towards the bead attachment site, as judged by reorientation of the microtubule-organizing center (MTOC) and localized actin polymerization. Thus, these cytoskeletal changes did not depend on activation of additional coreceptors. Moreover, single subunits of the TCR complex, namely TCR-zeta and CD3epsilon, were equally effective in inducing cytoskeletal polarization. However, mutagenesis of the immunoreceptor tyrosine-based activation motifs (ITAMs), present three times in TCR-zeta and once in CD3epsilon, revealed that the induction of cytoskeletal rearrangements required the presence of at least one intact ITAM. In agreement with this result, lack of functional Lck, the protein tyrosine kinase responsible for ITAM phosphorylation, abolished both MTOC reorientation and polarized actin polymerization. Both inhibitor and transient overexpression studies demonstrated that MTOC reorientation could occur in the absence of Ras activation. Our results suggest that APC-induced T cell polarization is a TCR-mediated event that is coupled to the TCR by the same signaling motif as TCR-induced gene activation, but diverges in its distal signaling requirements.
Figures
References
-
- Alberola-Ila J, Forbush KA, Seger R, Krebs EG, Perlmutter RM. Selective requirement for MAP kinase activation in thymocyte differentiation. Nature. 1995;373:620–623. - PubMed
-
- Anel A, Richieri GV, Kleinfeld AM. A tyrosine phosphorylation requirement for cytotoxic T lymphocyte degranulation. J Biol Chem. 1994;269:9506–9513. - PubMed
-
- Arpaia E, Shahar M, Dadi H, Cohen A, Roifman CM. Defective T cell receptor signaling and CD8+thymic selection in humans lacking ZAP-70 kinase. Cell. 1994;76:947–958. - PubMed
-
- Cardine AM, Kirchgessner H, Eckerskorn C, Meuer SC, Schraven B. Human T lymphocyte activation induces tyrosine phosphorylation of α-tubulin and its association with the SH2 domain of the p59fyn protein tyrosine kinase. Eur J Immunol. 1995;25:3290–3297. - PubMed
-
- Chan AC, Kadlecek TA, Elder ME, Filipovich AH, Kuo W-L, Iwashima M, Parslow TG, Weiss A. ZAP-70 deficiency in an autosomal recessive form of severe combined immunodeficiency. Science. 1994;264:1599–1601. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
