

Vascular endothelial growth factor induces endothelial fenestrations in vitro
- PMID: 9472045
- PMCID: PMC2141756
- DOI: 10.1083/jcb.140.4.947
Vascular endothelial growth factor induces endothelial fenestrations in vitro
Abstract
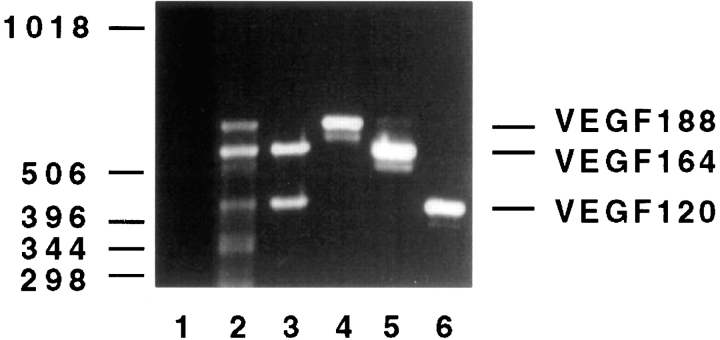
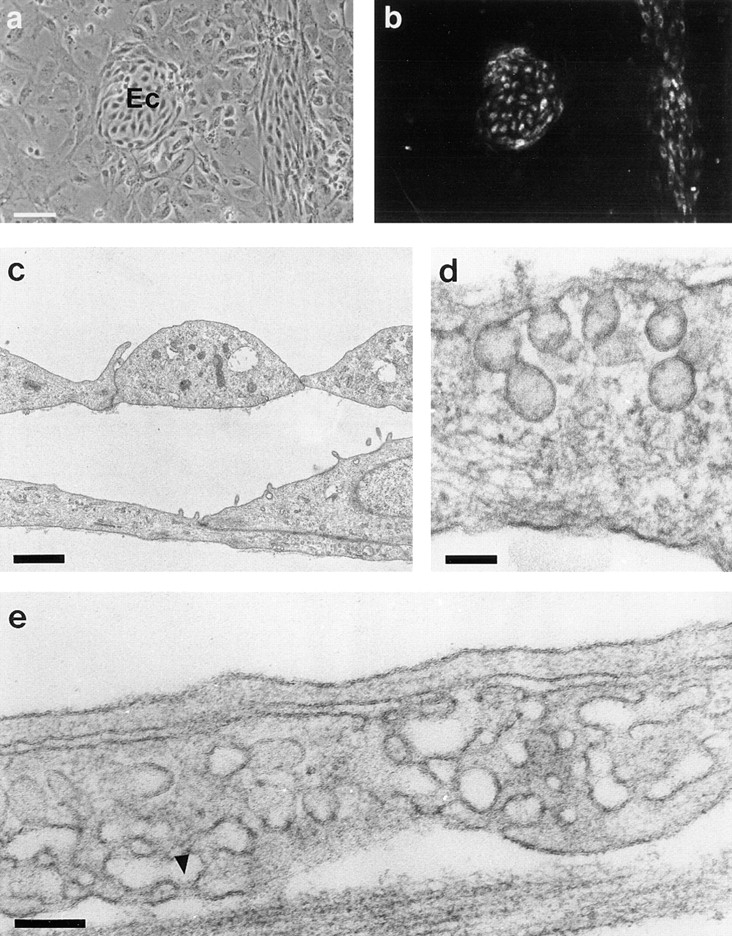
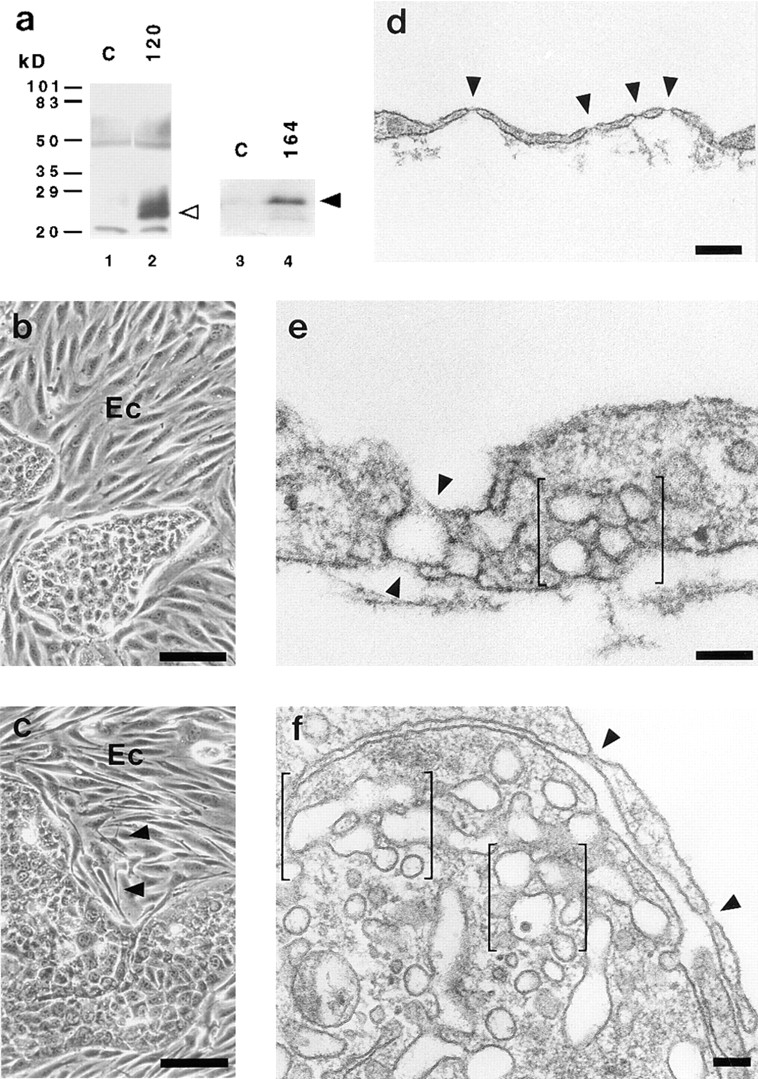
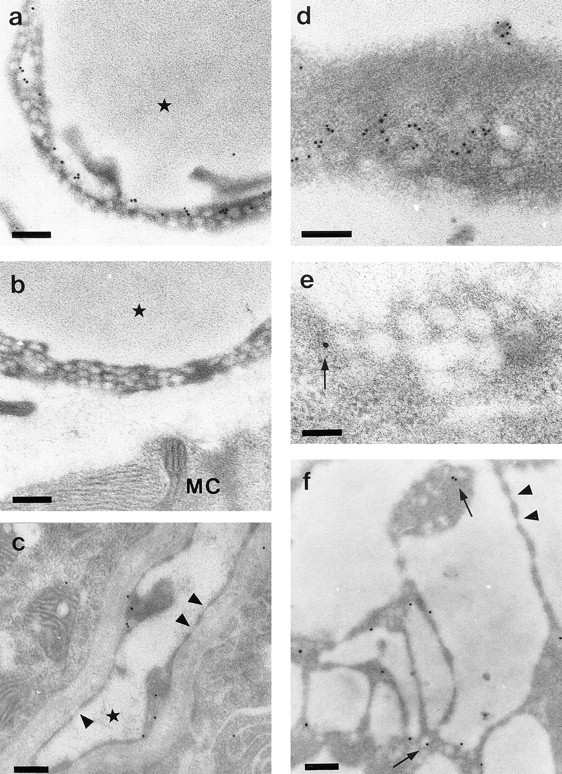
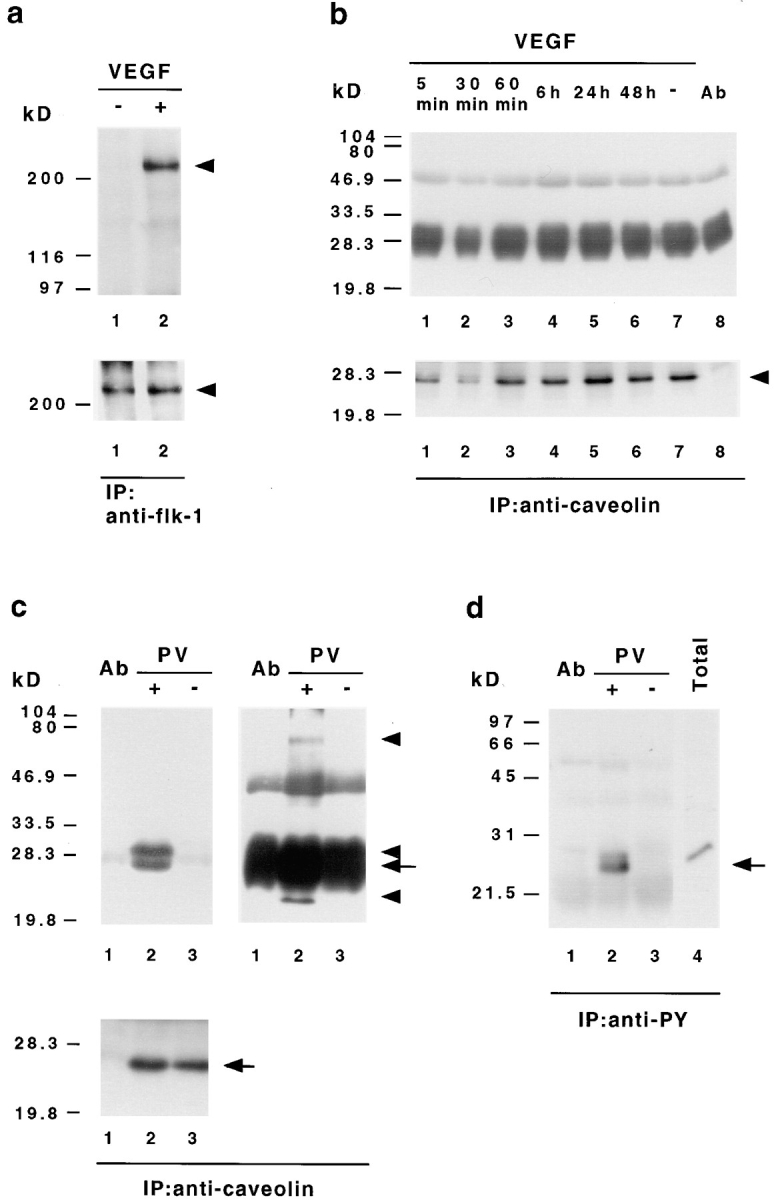
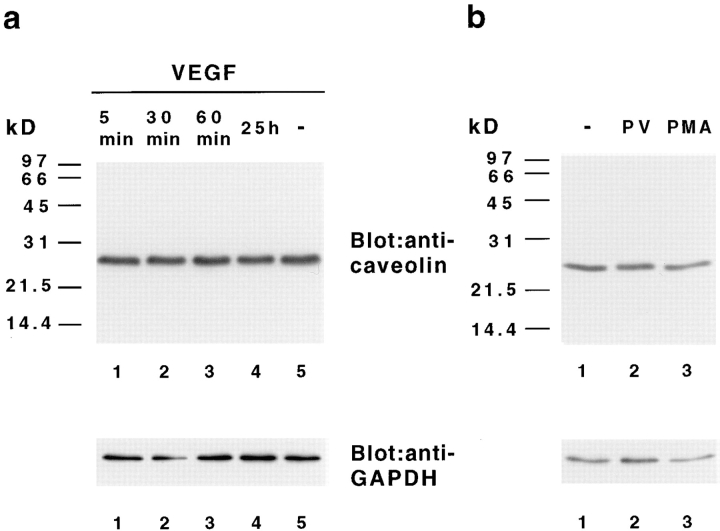
Vascular endothelial growth factor (VEGF) is an important regulator of vasculogenesis, angiogenesis, and vascular permeability. In contrast to its transient expression during the formation of new blood vessels, VEGF and its receptors are continuously and highly expressed in some adult tissues, such as the kidney glomerulus and choroid plexus. This suggests that VEGF produced by the epithelial cells of these tissues might be involved in the induction or maintenance of fenestrations in adjacent endothelial cells expressing the VEGF receptors. Here we describe a defined in vitro culture system where fenestrae formation was induced in adrenal cortex capillary endothelial cells by VEGF, but not by fibroblast growth factor. A strong induction of endothelial fenestrations was observed in cocultures of endothelial cells with choroid plexus epithelial cells, or mammary epithelial cells stably transfected with cDNAs for VEGF 120 or 164, but not with untransfected cells. These results demonstrate that, in these cocultures, VEGF is sufficient to induce fenestrations in vitro. Identical results were achieved when the epithelial cells were replaced by an epithelial-derived basal lamina-type extracellular matrix, but not with collagen alone. In this defined system, VEGF-mediated induction of fenestrae was always accompanied by an increase in the number of fused diaphragmed caveolae-like vesicles. Caveolae, but not fenestrae, were labeled with a caveolin-1-specific antibody both in vivo and in vitro. VEGF stimulation led to VEGF receptor tyrosine phosphorylation, but no change in the distribution, phosphorylation, or protein level of caveolin-1 was observed. We conclude that VEGF in the presence of a basal lamina-type extracellular matrix specifically induces fenestrations in endothelial cells. This defined in vitro system will allow further study of the signaling mechanisms involved in fenestrae formation, modification of caveolae, and vascular permeability.
Figures

References
-
- Bacic M, Edwards NA, Merrill MJ. Differential expression of vascular endothelial growth factor (vascular permeability factor) forms in rat tissues. Growth Factors. 1995;12:11–15. - PubMed
-
- Breier G, Albrecht U, Sterrer S, Risau W. Expression of vascular endothelial growth factor during embryonic angiogenesis and endothelial cell differentiation. Development (Camb) 1992;114:521–532. - PubMed
-
- Breier G, Clauss M, Risau W. Coordinate expression of vascular endothelial growth factor receptor-1 (flt-1) and its ligand suggests a paracrine regulation of murine vascular development. Dev Dyn. 1995;204:228–239. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
