

Molecular characterization of a phage-inducible middle promoter and its transcriptional activator from the lactococcal bacteriophage phi31
- PMID: 9473048
- PMCID: PMC106973
- DOI: 10.1128/JB.180.4.921-931.1998
Molecular characterization of a phage-inducible middle promoter and its transcriptional activator from the lactococcal bacteriophage phi31
Abstract
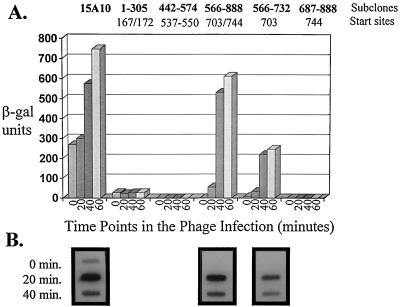
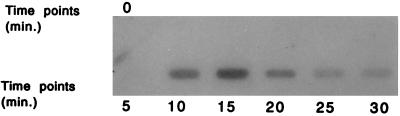
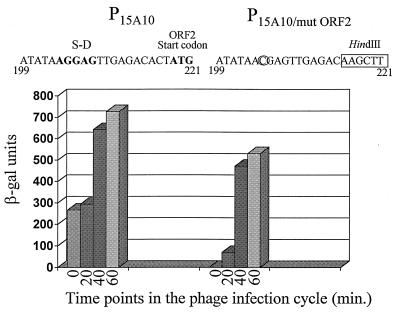
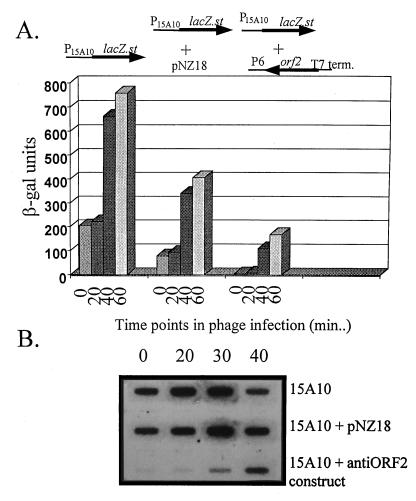
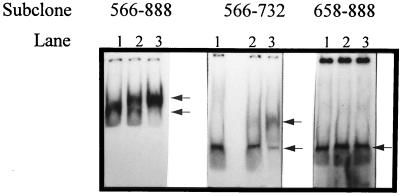
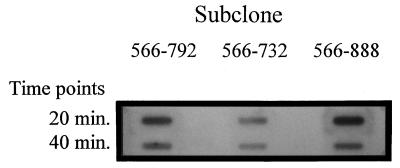
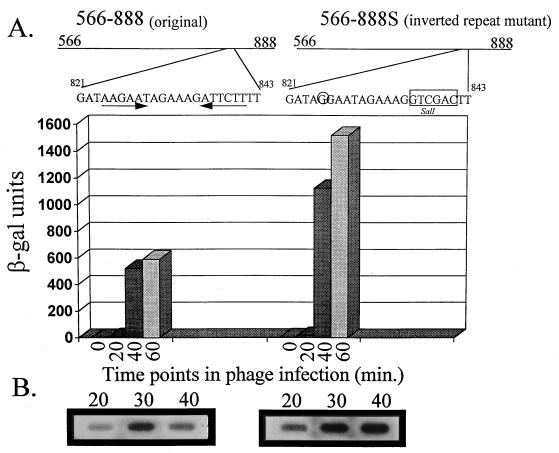
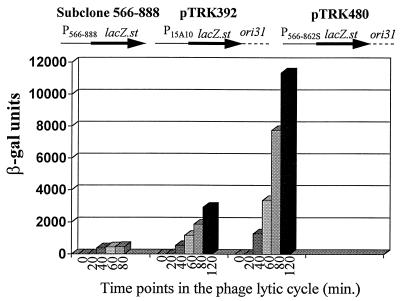
An inducible middle promoter from the lactococcal bacteriophage phi31 was isolated previously by shotgun cloning an 888-bp fragment (P15A10) upstream of the beta-galactosidase (beta-Gal) gene (lacZ.st) from Streptococcus thermophilus (D. J. O'Sullivan, S. A. Walker, S. G. West, and T. R. Klaenhammer, Bio/Technology 14:82-87, 1996). The promoter showed low levels of constitutive beta-Gal activity which could be induced two- to threefold over baseline levels after phage infection. During this study, the fragment was subcloned and characterized to identify a smaller, tightly regulated promoter fragment which allowed no beta-Gal activity until after phage infection. This fragment, defined within nucleotides 566 to 888 (P(566-888); also called fragment 566-888), contained tandem, phage-inducible transcription start sites at nucleotides 703 and 744 (703/744 start sites). Consensus -10 regions were present upstream of both start sites, but no consensus -35 regions were identified for either start site. A transcriptional activator, encoded by an open reading frame (ORF2) upstream of the 703/744 start sites, was identified for P(566-888). ORF2 activated P(566-888) when provided in trans in Escherichia coli. In addition, when combined with pTRK391 (P15A10::lacZ.st) in Lactococcus lactis NCK203, an antisense ORF2 construct was able to retard induction of the phage-inducible promoter as measured by beta-Gal activity levels. Finally, gel shift assays showed that ORF2 was able to bind to promoter fragment 566-888. Deletion analysis of the region upstream from the tandem promoters identified a possible binding site for transcriptional activation of the phage promoters. The DNA-binding ability of ORF2 was eliminated upon deletion of part of this region, which lies centered approximately 35 bp upstream of start site 703. Deletion analysis and mutagenesis studies also elucidated a critical region downstream of the 703/744 start sites, where mutagenesis resulted in a two- to threefold increase in beta-Gal activity. With these improvements, the level of expression achieved by an explosive-expression strategy was elevated from 3,000 to 11,000 beta-Gal units within 120 min after induction.
Figures



Similar articles
-
Common elements regulating gene expression in temperate and lytic bacteriophages of Lactococcus species.Appl Environ Microbiol. 1998 Mar;64(3):1147-52. doi: 10.1128/AEM.64.3.1147-1152.1998. Appl Environ Microbiol. 1998. PMID: 9501453 Free PMC article.
-
Molecular characterization of a genomic region in a Lactococcus bacteriophage that is involved in its sensitivity to the phage defense mechanism AbiA.J Bacteriol. 1997 May;179(9):2949-57. doi: 10.1128/jb.179.9.2949-2957.1997. J Bacteriol. 1997. PMID: 9139913 Free PMC article.
-
An explosive antisense RNA strategy for inhibition of a lactococcal bacteriophage.Appl Environ Microbiol. 2000 Jan;66(1):310-9. doi: 10.1128/AEM.66.1.310-319.2000. Appl Environ Microbiol. 2000. PMID: 10618241 Free PMC article.
-
Use of the integration elements encoded by the temperate lactococcal bacteriophage TP901-1 to obtain chromosomal single-copy transcriptional fusions in Lactococcus lactis.Appl Environ Microbiol. 1999 Feb;65(2):752-8. doi: 10.1128/AEM.65.2.752-758.1999. Appl Environ Microbiol. 1999. PMID: 9925612 Free PMC article.
-
Manufacturing Bacteriophages (Part 1 of 2): Cell Line Development, Upstream, and Downstream Considerations.Pharmaceuticals (Basel). 2021 Sep 17;14(9):934. doi: 10.3390/ph14090934. Pharmaceuticals (Basel). 2021. PMID: 34577634 Free PMC article. Review.
Cited by
-
Lactococcus lactis lytic bacteriophages of the P335 group are inhibited by overexpression of a truncated CI repressor.J Bacteriol. 2002 Dec;184(23):6532-44. doi: 10.1128/JB.184.23.6532-6543.2002. J Bacteriol. 2002. PMID: 12426341 Free PMC article.
-
Common elements regulating gene expression in temperate and lytic bacteriophages of Lactococcus species.Appl Environ Microbiol. 1998 Mar;64(3):1147-52. doi: 10.1128/AEM.64.3.1147-1152.1998. Appl Environ Microbiol. 1998. PMID: 9501453 Free PMC article.
-
Temporal transcription map of the virulent Streptococcus thermophilus bacteriophage Sfi19.Appl Environ Microbiol. 2004 Aug;70(8):5041-6. doi: 10.1128/AEM.70.8.5041-5046.2004. Appl Environ Microbiol. 2004. PMID: 15294848 Free PMC article.
-
Analysis of the genetic switch and replication region of a P335-type bacteriophage with an obligate lytic lifestyle on Lactococcus lactis.Appl Environ Microbiol. 2001 Mar;67(3):1128-39. doi: 10.1128/AEM.67.3.1128-1139.2001. Appl Environ Microbiol. 2001. PMID: 11229902 Free PMC article.
-
Transcription analysis of the prolate-headed lactococcal bacteriophage c2.J Bacteriol. 1998 Sep;180(17):4487-96. doi: 10.1128/JB.180.17.4487-4496.1998. J Bacteriol. 1998. PMID: 9721287 Free PMC article.
References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Casabadan M J, Cohen S N. Analysis of gene control signals by DNA fusion and cloning in Escherichia coli. J Mol Biol. 1980;138:179–207. - PubMed
-
- Chung D K, Chung S K, Batt C A. Antisense RNA directed against the major capsid protein of Lactococcus lactis subsp. cremoris bacteriophage F4-1 confers partial resistance to the host. Appl Microbiol Biotechnol. 1992;37:79–83. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
