

Inhibition by ATP of hippocampal synaptic transmission requires localized extracellular catabolism by ecto-nucleotidases into adenosine and channeling to adenosine A1 receptors
- PMID: 9482785
- PMCID: PMC6792930
- DOI: 10.1523/JNEUROSCI.18-06-01987.1998
Inhibition by ATP of hippocampal synaptic transmission requires localized extracellular catabolism by ecto-nucleotidases into adenosine and channeling to adenosine A1 receptors
Abstract
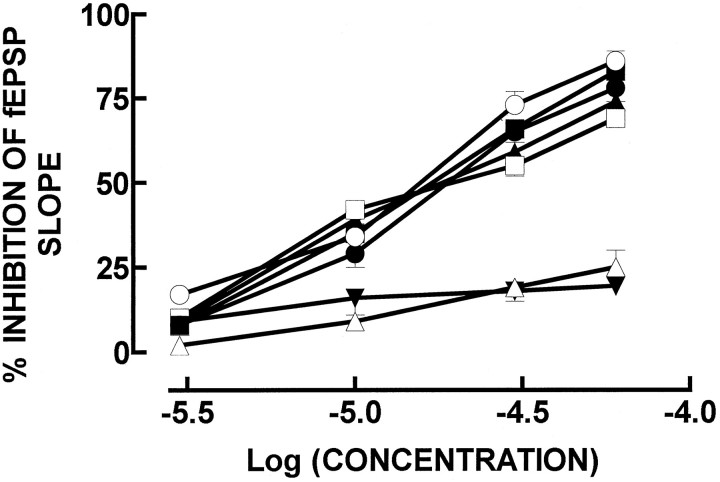
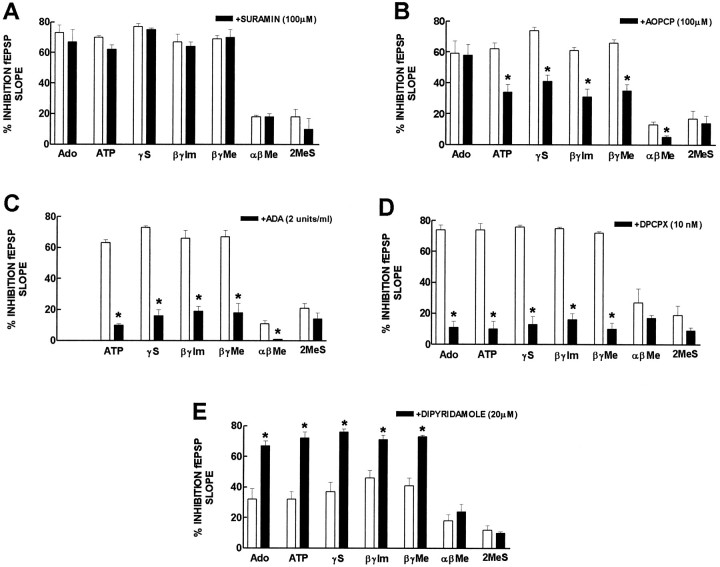
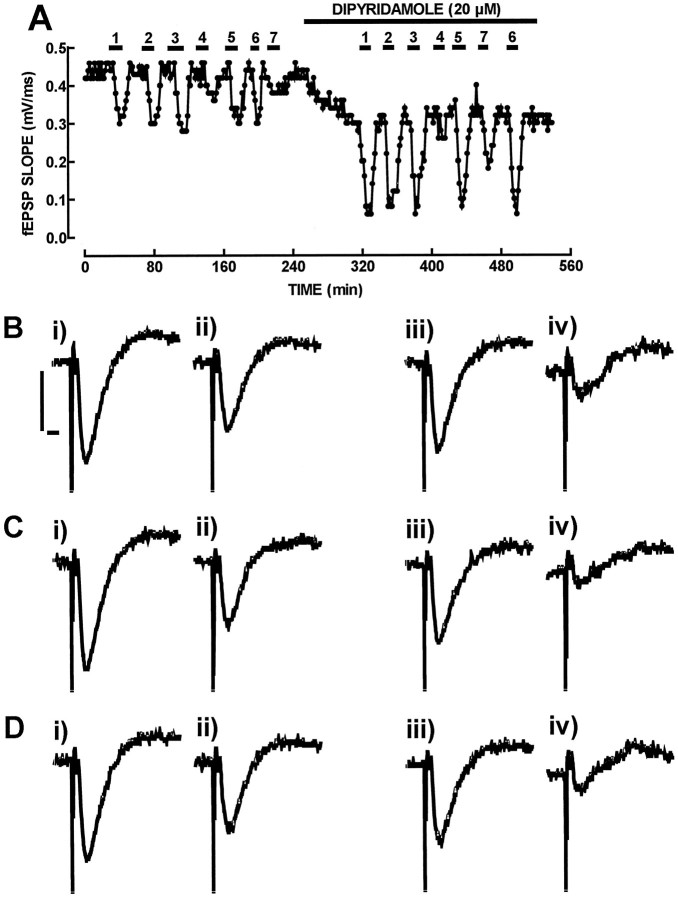
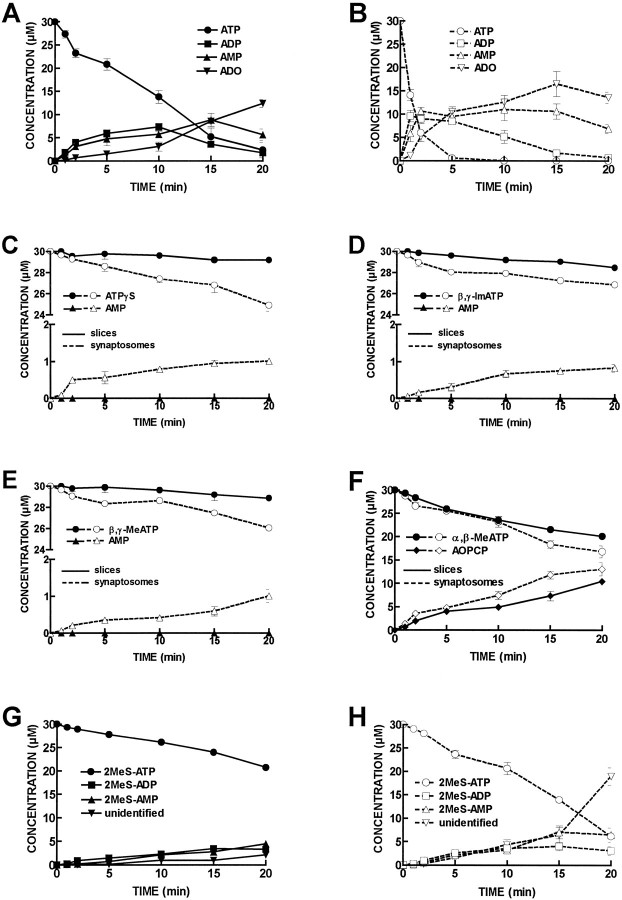
ATP analogs substituted in the gamma-phosphorus (ATPgammaS, beta, gamma-imido-ATP, and beta,gamma-methylene-ATP) were used to probe the involvement of P2 receptors in the modulation of synaptic transmission in the hippocampus, because their extracellular catabolism was virtually not detected in CA1 slices. ATP and gamma-substituted analogs were equipotent to inhibit synaptic transmission in CA1 pyramid synapses (IC50 of 17-22 microM). The inhibitory effect of ATP and gamma-phosphorus-substituted ATP analogs (30 microM) was not modified by the P2 receptor antagonist suramin (100 microM), was inhibited by 42-49% by the ecto-5'-nucleotidase inhibitor and alpha,beta-methylene ADP (100 microM), was inhibited by 74-85% by 2 U/ml adenosine deaminase (which converts adenosine into its inactive metabolite-inosine), and was nearly prevented by the adenosine A1 receptor antagonist 1,3-dipropyl-8-cyclopentylxanthine (10 nM). Stronger support for the involvement of extracellular adenosine formation as a main requirement for the inhibitory effect of ATP and gamma-substituted ATP analogs was the observation that an inhibitor of adenosine uptake, dipyridamole (20 microM), potentiated by 92-124% the inhibitory effect of ATP and gamma-substituted ATP analogs (10 microM), a potentiation similar to that obtained for 10 microM adenosine (113%). Thus, the present results indicate that inhibition by extracellular ATP of hippocampal synaptic transmission requires localized extracellular catabolism by ecto-nucleotidases and channeling of the generated adenosine to adenosine A1 receptors.
Figures
References
-
- Bruns RF. Adenosine receptors. Roles and pharmacology. Ann NY Acad Sci. 1990;603:211–226. - PubMed
-
- Cascalheira JF, Sebastião AM. Adenine nucleotide analogues, including γ-phosphate-substituted analogues, are metabolised extracellularly in innervated frog sartorius muscle. Eur J Pharmacol. 1992;222:49–59. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous