

Brome mosaic virus RNA replication protein 1a dramatically increases in vivo stability but not translation of viral genomic RNA3
- PMID: 9482867
- PMCID: PMC19301
- DOI: 10.1073/pnas.95.5.2227
Brome mosaic virus RNA replication protein 1a dramatically increases in vivo stability but not translation of viral genomic RNA3
Abstract
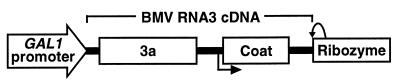
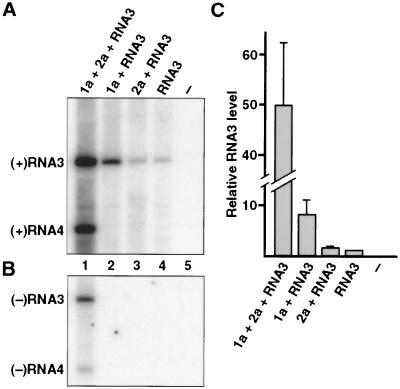
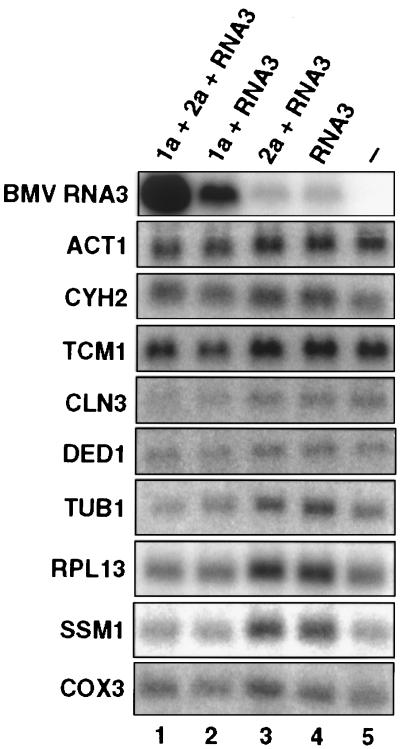
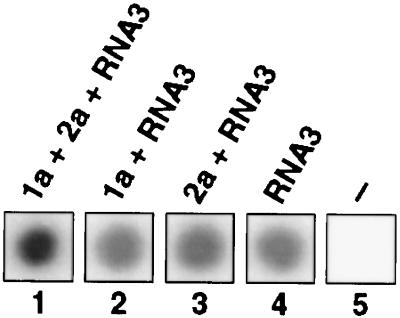
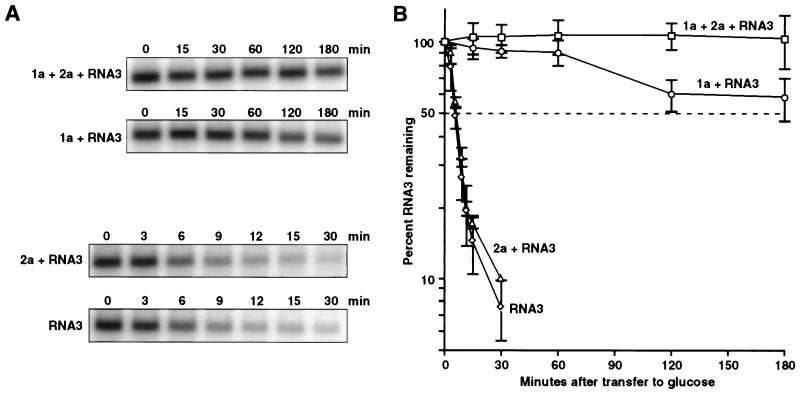
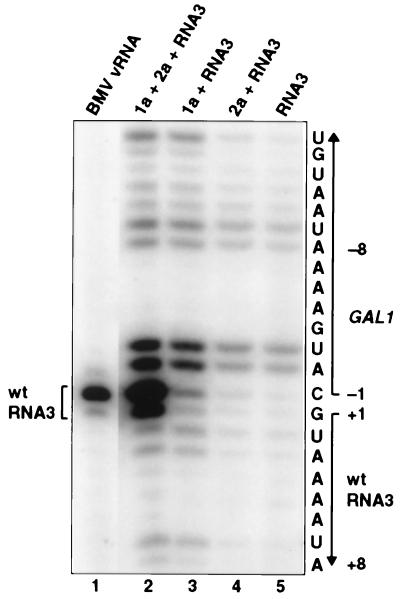
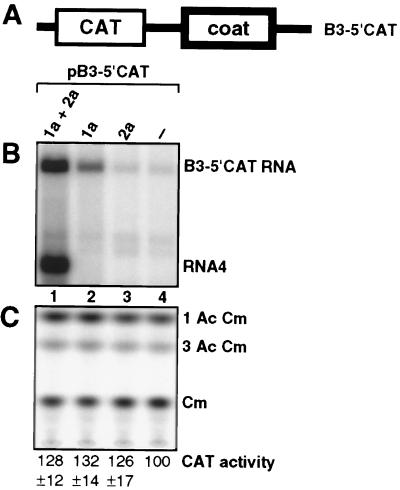
Brome mosaic virus (BMV), a positive-strand RNA virus in the alphavirus-like superfamily, encodes two RNA replication proteins: 1a, which contains a helicase-like domain and a domain implicated in RNA capping, and 2a, which contains a polymerase-like domain. To further explore their functions, we expressed 1a and 2a individually and together in yeast also expressing replicatable transcripts of BMV genomic RNA3. Complementing prior results that 1a and 2a are required jointly for positive-strand RNA synthesis, both also were required for negative-strand RNA synthesis. Nevertheless, in the absence of 2a, 1a expression increased the accumulation of DNA-derived RNA3 transcripts 8-fold. Increased accumulation was specific for RNA3: none of a diverse set of yeast mRNAs tested showed increased accumulation in the presence of 1a. Increased RNA3 accumulation was not due to increased DNA transcription, but to a 20- to 40-fold increase in the in vivo half-life of RNA3 from 5-10 min in the absence of 1a to more than 3 hr in the presence of 1a. Although (1a+2a)-dependent RNA replication selectively amplified the natural viral 5' end from among multiple transcription starts of DNA-derived RNA3 transcripts, 1a-induced stabilization affected all RNA3 transcripts, without specificity for the precise viral 5' end. Increased RNA3 accumulation did not increase expression of a directly translatable, 5'-proximal gene in RNA3, implying that 1a-induced stabilization blocked rather than facilitated RNA3 translation. These and other results suggest that the striking, 1a-induced stabilization of RNA3 may reflect an interaction involved in recruiting viral RNA templates into RNA replication while diverting them from the competing processes of translation and degradation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
