

RNA-binding protein TIAR is essential for primordial germ cell development
- PMID: 9482885
- PMCID: PMC19335
- DOI: 10.1073/pnas.95.5.2331
RNA-binding protein TIAR is essential for primordial germ cell development
Abstract
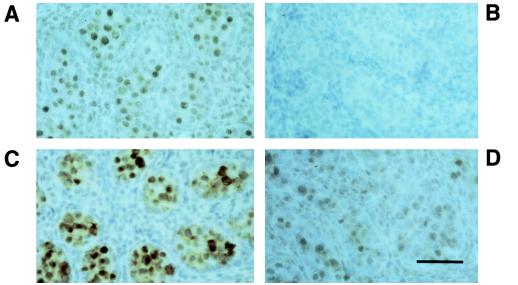
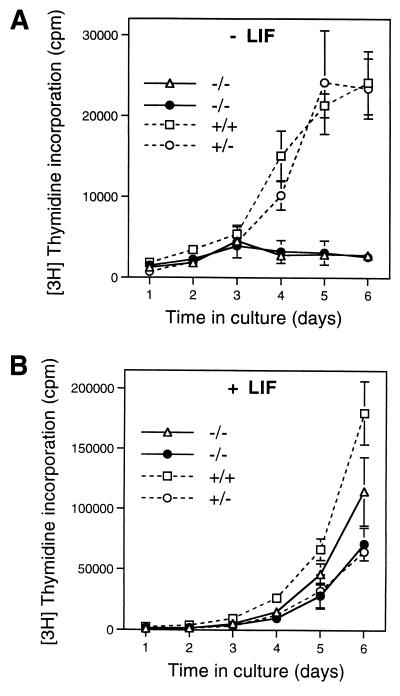
Primordial germ cells (PGCs) give rise to both eggs and sperm via complex maturational processes that require both cell migration and proliferation. However, little is known about the genes controlling gamete formation during the early stages of PGC development. Although several mutations are known to severely reduce the number of PGCs reaching and populating the genital ridges, the molecular identity of only two of these genes is known: the c-kit receptor protein tyrosine kinase and the c-kit ligand (the steel factor). Herein, we report that mutant mice lacking TIAR, an RNA recognition motif/ribonucleoprotein-type RNA-binding protein highly expressed in PGCs, fail to develop spermatogonia or oogonia. This developmental defect is a consequence of reduced survival of PGCs that migrate to the genital ridge around embryonic day 11.5 (E11.5). The numbers of PGCs populating the genital ridge in TIAR-deficient embryos are severely reduced compared to wild-type embryos by E11.5 and in the mutants PGCs are completely absent at E13.5. Furthermore, TIAR-deficient embryonic stem cells do not proliferate in the absence of exogenous leukemia inhibitory factor in an in vitro methylcellulose culture assay, supporting a role for TIAR in regulating cell proliferation. Because the development of PGCs relies on the action of several growth factors, these results are consistent with a role for TIAR in the expression of a survival factor or survival factor receptor that is essential for PGC development. TIAR-deficient mice thus provide a model system to study molecular mechanisms of PGC development and possibly the basis for some forms of idiopathic infertility.
Figures
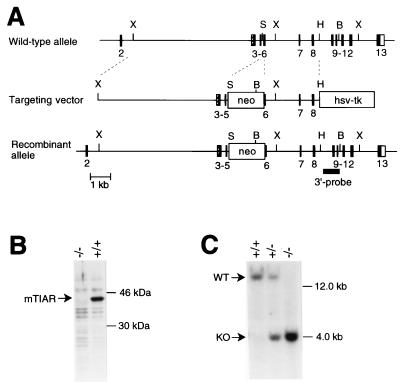
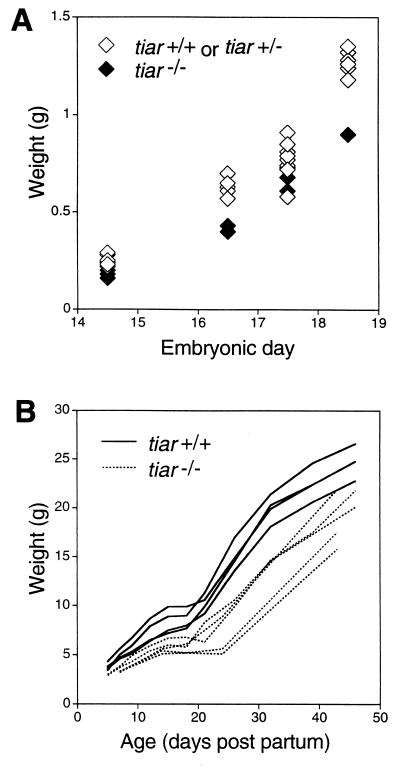
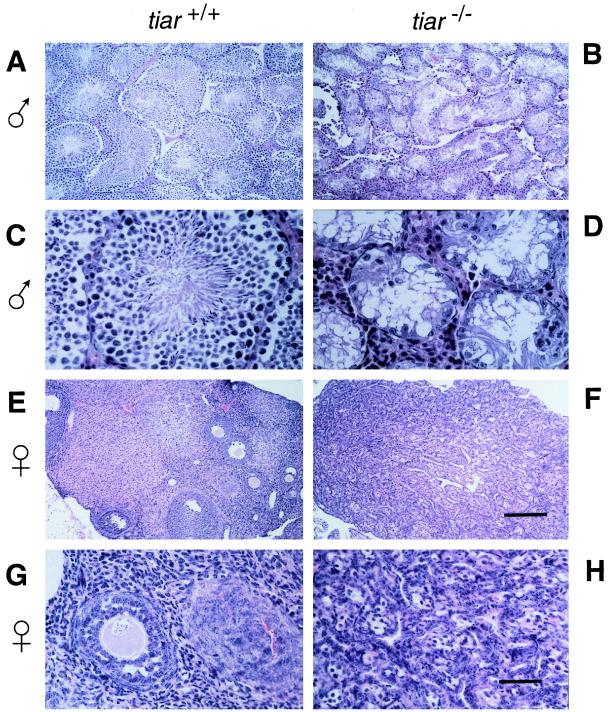
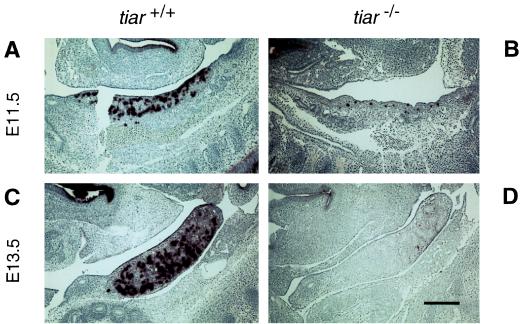
References
-
- Hogan B, Beddington R, Costantini F, Lacy E. Manipulating the Mouse Embryo: A Laboratory Manual. Plainview, NY: Cold Spring Harbor Lab. Press; 1994.
-
- McLaren A. In: Organization of the Early Vertebrate Embryo. Zagris N, Duprat A M, Durston A J, editors. New York: Plenum; 1995. pp. 1–9.
-
- Lawson K A, Hage W J. CIBA Found Symp. 1994;182:68–84. - PubMed
-
- Ginsburg M, Snow M H L, McLaren A. Development (Cambridge, UK) 1990;110:521–528. - PubMed
-
- Eddy E M, Clark J M, Gong D, Fenderson B A. Gamete Res. 1981;4:333–362.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
