

Fatty acids suppress voltage-gated Na+ currents in HEK293t cells transfected with the alpha-subunit of the human cardiac Na+ channel
- PMID: 9482947
- PMCID: PMC19460
- DOI: 10.1073/pnas.95.5.2680
Fatty acids suppress voltage-gated Na+ currents in HEK293t cells transfected with the alpha-subunit of the human cardiac Na+ channel
Abstract
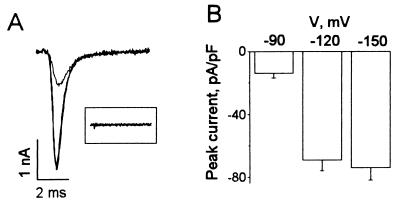
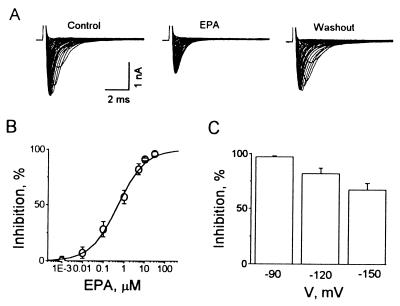
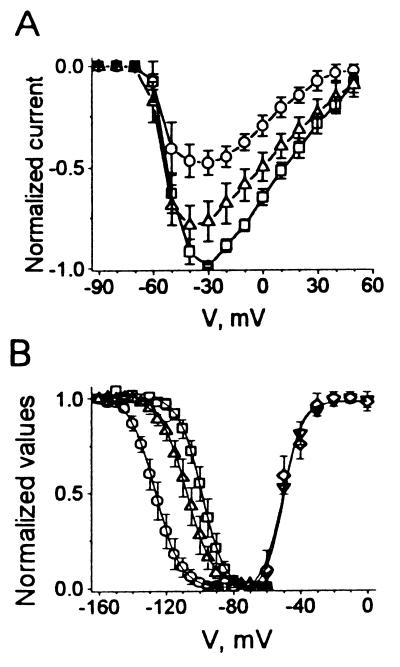
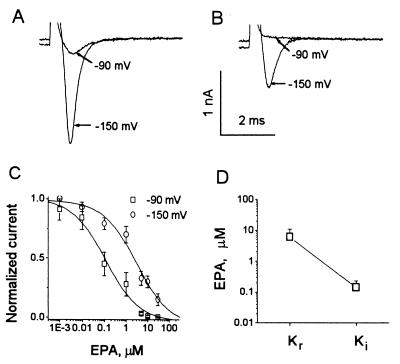
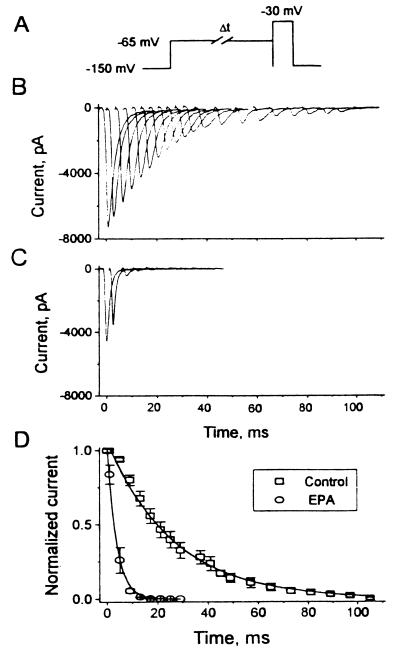
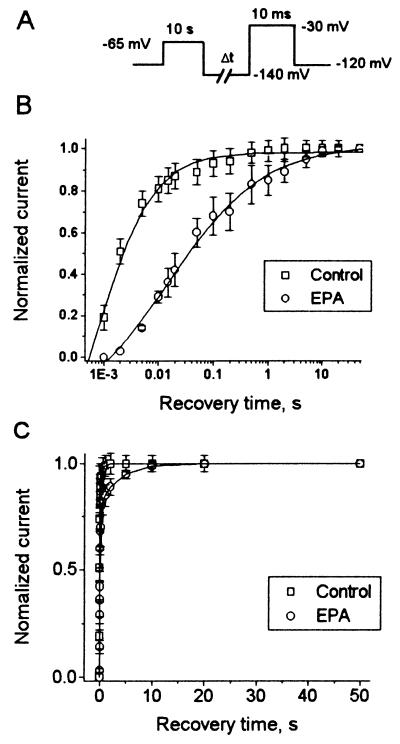
Studies have shown that fish oils, containing n-3 fatty acids, have protective effects against ischemia-induced, fatal cardiac arrhythmias in animals and perhaps in humans. In this study we used the whole-cell voltage-clamp technique to assess the effects of dietary, free long-chain fatty acids on the Na+ current (INa,alpha) in human embryonic kidney (HEK293t) cells transfected with the alpha-subunit of the human cardiac Na+ channel (hH1alpha). Extracellular application of 0.01 to 30 microM eicosapentaenoic acid (EPA, C20:5n-3) significantly reduced INa,alpha with an IC50 of 0.51 +/- 0.06 microM. The EPA-induced suppression of INa,alpha was concentration- and voltage-dependent. EPA at 5 microM significantly shifted the steady-state inactivation relationship by -27.8 +/- 1.2 mV (n = 6, P < 0.0001) at the V1/2 point. In addition, EPA blocked INa,alpha with a higher "binding affinity" to hH1alpha channels in the inactivated state than in the resting state. The transition from the resting state to the inactivated state was markedly accelerated in the presence of 5 microM EPA. The time for 50% recovery from the inactivation state was significantly slower in the presence of 5 microM EPA, from 2.1 +/- 0.8 ms for control to 34.8 +/- 2.1 ms (n = 5, P < 0.001). The effects of EPA on INa,alpha were reversible. Furthermore, docosahexaenoic acid (C22:6n-3), alpha-linolenic acid (C18:3n-3), conjugated linoleic acid (C18:2n-7), and oleic acid (C18:1n-9) at 5 microM and all-trans-retinoic acid at 10 microM had similar effects on INa,alpha as EPA. Even 5 microM of stearic acid (C18:0) or palmitic acid (C16:0) also significantly inhibited INa, alpha. In contrast, 5 microM EPA ethyl ester did not alter INa,alpha (8 +/- 4%, n = 8, P > 0.05). The present data demonstrate that free fatty acids suppress INa,alpha with high "binding affinity" to hH1alpha channels in the inactivated state and prolong the duration of recovery from inactivation.
Figures
References
-
- McLennan P L, Abeywardena M Y, Charnock J S. Can J Physiol Pharmacol. 1985;63:1411–1417. - PubMed
-
- McLennan P L, Bridle T M, Abeywardena M Y, Charnock J S. Am Heart J. 1992;123:1555–1561. - PubMed
-
- Hock C E, Beck L D, Bodine L C, Reibel D K. Am J Physiol. 1990;259:H1518–H1526. - PubMed
-
- Burr M, Gilbert J F, Holliday R M, Elwood P C, Fehily A M, Rogers S, Sweetnam P M, Deadman N M. Lancet. 1989;334:757–761. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
