

doi: 10.1128/MCB.18.3.1156.
Nucleosome translational position, not histone acetylation, determines TFIIIA binding to nucleosomal Xenopus laevis 5S rRNA genes
Affiliations
- PMID: 9488430
- PMCID: PMC108828
- DOI: 10.1128/MCB.18.3.1156
Item in Clipboard
Nucleosome translational position, not histone acetylation, determines TFIIIA binding to nucleosomal Xenopus laevis 5S rRNA genes
Mol Cell Biol.
1998 Mar.
Abstract
We sought to study the binding constraints placed on the nine-zinc-finger protein transcription factor IIIA (TFIIIA) by a histone octamer. To this end, five overlapping fragments of the Xenopus laevis oocyte and somatic 5S rRNA genes were reconstituted into nucleosomes, and it was subsequently shown that nucleosome translational positioning is a major determinant of the binding of TFIIIA to the 5S rRNA genes. Furthermore, it was found that histone acetylation cannot override the TFIIIA binding constraints imposed by unfavorable translational positions.
Figures
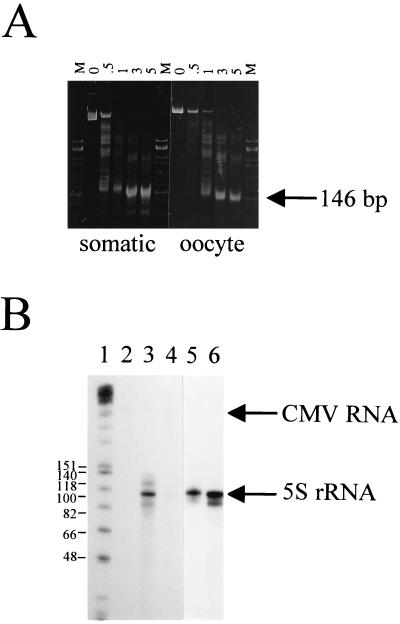
Effect of nucleosome reconstitution on 5S rRNA gene transcription. (A) Micrococcal nuclease digestion of nucleosomes reconstituted on full-length X. laevis oocyte and somatic genes. Digestions were carried out at a nucleosome concentration of 0.1 mg/ml (DNA weight) and an enzyme concentration of 10 U/ml for the times (minutes) indicated above each lane. The resulting DNA fragments were deproteinized and electrophoresed on a 4% nondenaturing gel. Lanes M, HhaI-cut pBR322. (B) Approximately 250 ng of the oocyte (lanes 3 and 4) or somatic (lanes 5 and 6) 5S rRNA genes, either uncomplexed (lanes 3 and 5) or reconstituted with histones isolated from chicken erythrocytes (lanes 4 and 6), was transcribed in HeLa cell nuclear extracts supplemented with 150 nM recombinant Xenopus TFIIIA and 2 mM MgCl2. Each reaction mixture contained 50 ng of CMV DNA as an internal transcription control, which is visible only after longer exposures. Transcripts were analyzed by denaturing polyacrylamide gel electrophoresis (8% acrylamide and 8.3 M urea in 1× TBE). Lane 1, Klenow fragment-end-labeled HinfI-cut φX174 DNA (sizes of marker fragments are shown as numbers of nucleotides); lane 2, transcribed CMV DNA alone.
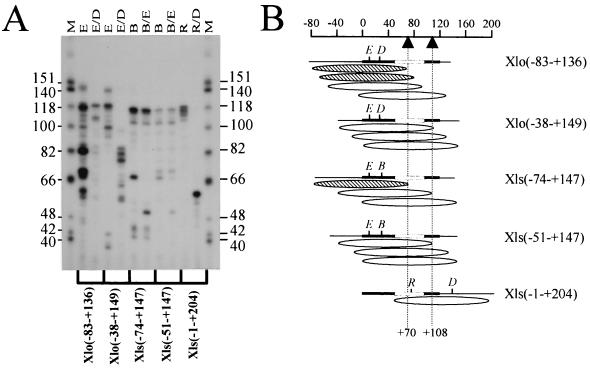
Determination of nucleosome translational position on reconstituted X. laevis 5S rRNA gene fragments. (A) The region of DNA in direct association with the histone octamer was determined by digestion of an approximately 146-bp, internally labeled, micrococcal nuclease-resistant fragment with different combinations of restriction enzymes as indicated. The resulting restriction fragments were resolved by denaturing polyacrylamide gel electrophoresis (8% acrylamide and 8.3 M urea in 1× TBE). Lanes M, HinfI-cut φX174 DNA used as a marker (sizes of marker fragments are shown as numbers of nucleotides). The restriction enzymes used are indicated above the gel: E, EaeI; D, DdeI; B, Bsp1286; R, RsaI. (B) Schematic representation of the most predominant nucleosome positions on the five different fragments of the X. laevis oocyte and somatic 5S rRNA genes resulting from the electrophoretic analysis shown in panel A. Due to variations in base composition and thus radioactive labeling, the intensities of the bands in relation to the dATP contents of the fragments were considered in these calculations. The ellipsoids indicate the most abundant positions of the approximately 146-bp micrococcal nuclease-resistant fragments. The thick black lines represent the 5S rRNA coding sequence, and the open boxes indicate the intragenic TFIIIA binding site. Nucleotide positions relative to the transcription start site are indicated on the scale at the top, and the dashed vertical lines indicate positions +70 and +108. The hatched ellipsoids are those nucleosome positions postulated to allow TFIIIA binding, whereas open ellipsoids are those which are thought to be repressive to TFIIIA binding based on the results of You et al. (48).
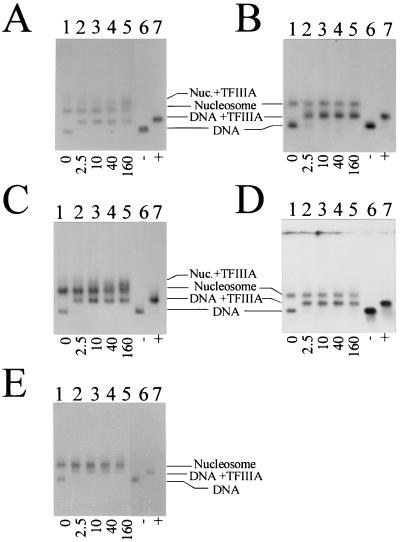
Analysis of the binding of TFIIIA to nucleosomes reconstituted onto the X. laevis 5S rRNA gene fragments by agarose gel electrophoresis. (A) Xlo(−83→+136); (B) Xlo(−38→+149); (C) Xls(−74→+147); (D) Xls(−51→+147); (E) Xls(−1→+204). Lanes 1 to 5, nucleosomes reconstituted with histones isolated from non-butyrate-treated HeLa cells in the absence (lane 1) or presence (lanes 2 to 5) of increasing amounts of TFIIIA (molar ratios are indicated below the gels). Lanes 6 and 7, corresponding DNA templates in the absence (−) or presence (+) of TFIIIA. The samples were incubated for 20 min at room temperature before being loaded on agarose gels.
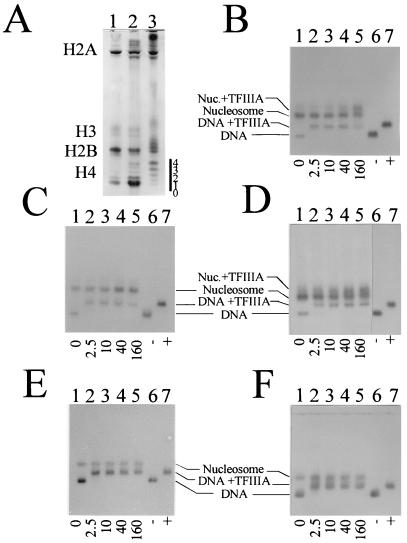
(A) Acetic acid (6%)–urea (8 M)–Triton X-100 (8 mM) electrophoretic analysis of the histones from the nucleosome core particles used in the exchange reconstitutions. Lane 1, chicken erythrocyte core particles; lane 2, HeLa cell nucleosome core particles; lane 3, nucleosome core particles (fraction A [4]) from butyrate-treated HeLa cells. The number of acetyl groups on histone H4 is indicated by the numbers to the right of the gel. (B to F) Analysis of the binding of TFIIIA to nucleosomes containing hyperacetylated HeLa histones. The five X. laevis 5S rRNA gene fragments were tested for binding after reconstitution with nucleosome core particles isolated from sodium butyrate-treated HeLa cells. (B) Xlo(−83→+136); (C) Xlo(−38→+149); (D) Xls(−74→+147); (E) Xls(−51→+147); (F) Xls(−1→+204). Lanes 1, no TFIIIA; lanes 2 to 5, increasing amounts of TFIIIA (molar ratios are indicated below the gels); lanes 6 and 7, corresponding DNA templates in the absence (−) or presence (+) of TFIIIA. The samples were incubated for 20 min at room temperature before being loaded on agarose gels.
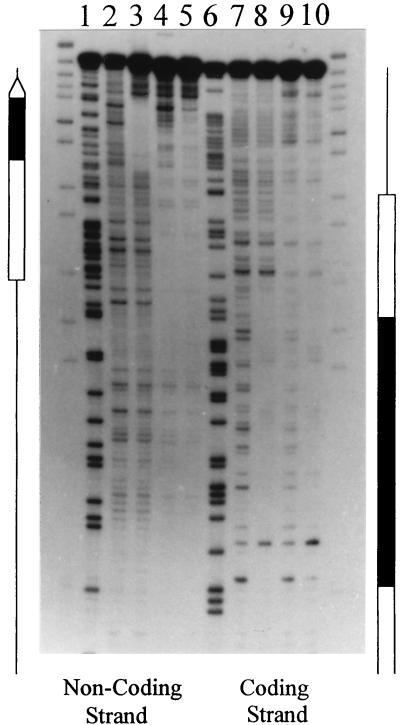
DNase I footprint analysis of the complexes formed by nucleosomes and/or TFIIIA on the Xlo(−83→+136) 5S rRNA gene fragment (Fig. 3B). Nucleosomes labeled at the 3′ end of the coding and noncoding strands were incubated in the presence of TFIIIA and subsequently digested with DNase I. The partially digested TFIIIA-nucleosome complexes were purified from free DNA and unbound nucleosomes by native gel electrophoresis (see Materials and Methods). The footprints for the coding and noncoding strands for naked DNA (lanes 2 and 7), TFIIIA-bound DNA (lanes 3 and 8), reconstituted nucleosome (lanes 4 and 9), and TFIIIA-nucleosome complexes (lanes 5 and 10) are shown. Also shown are the Maxam-Gilbert reactions of the labeled DNA (lanes 1 and 6). The 5S rRNA gene is indicated by the open arrow, and the TFIIIA binding site is indicated by a black box.
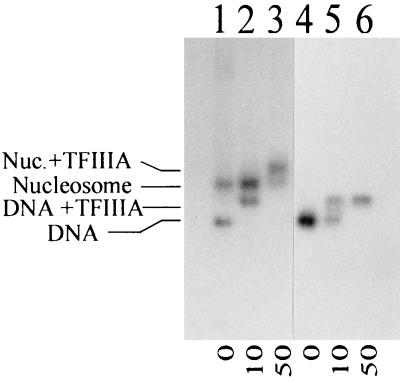
Analysis of the binding of TFIIIA to nucleosomes reconstituted onto the X. borealis 5S rRNA gene fragment by agarose gel electrophoresis, showing nucleosomes (lanes 1 to 3) or uncomplexed DNA (lanes 4 to 6) in the absence (lanes 1 and 4) or presence (lanes 2, 3, 5, and 6) of increasing amounts of TFIIIA. The nucleosomes used for the reconstitution were isolated from chicken erythrocytes, and the relative molar ratios of TFIIIA are indicated below the gel.
References
-
- Ausió J. Structure and dynamics of transcriptionally active chromatin. J Cell Sci. 1992;102:1–5. - PubMed
-
- Ausió J, Dong F, van Holde K E. Use of selectively trypsinized nucleosome core particles to analyze the role of the histone “tails” in the stabilization of the nucleosome. J Mol Biol. 1989;206:451–463. - PubMed
-
- Ausió J, Seger D S, Eisenberg H. Nucleosome core particle stability and conformational change. J Mol Biol. 1984;176:77–104. - PubMed
-
- Ausió J, van Holde K E. Histone hyperacetylation: its effects on nucleosome conformation and stability. Biochemistry. 1986;25:1421–1428. - PubMed
-
- Blomquist P, Li Q, Wrange O. The affinity of nuclear factor 1 for its DNA site is drastically reduced by nucleosome organization irrespective of its rotational or translational position. J Biol Chem. 1996;271:153–159. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources