

Architecture of protein and DNA contacts within the TFIIIB-DNA complex
- PMID: 9488485
- PMCID: PMC108883
- DOI: 10.1128/MCB.18.3.1682
Architecture of protein and DNA contacts within the TFIIIB-DNA complex
Abstract
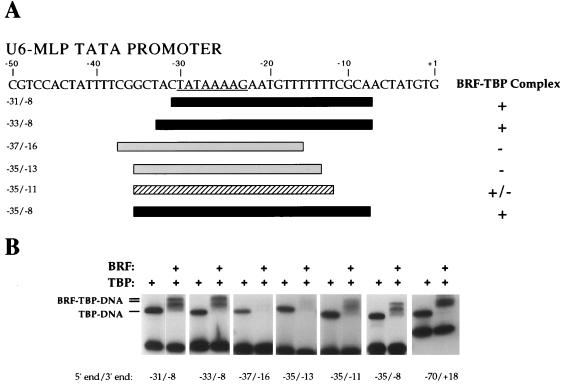
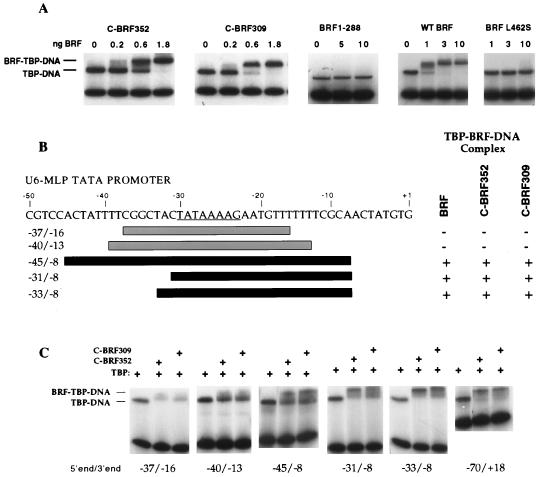
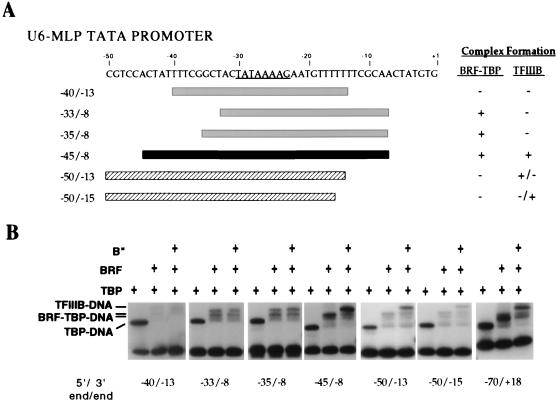
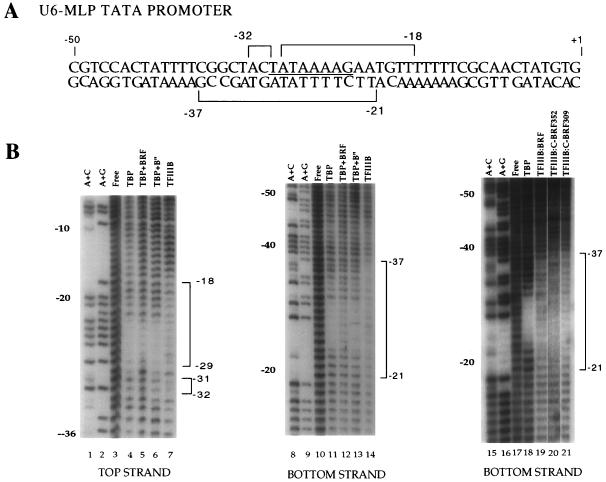
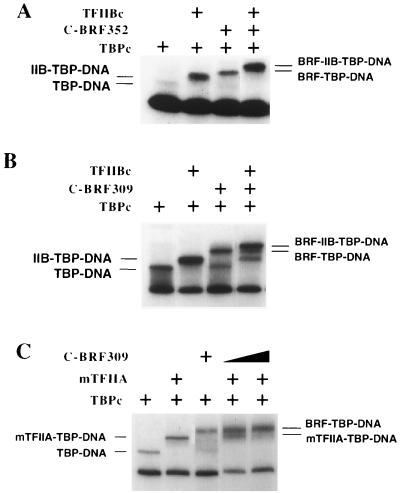
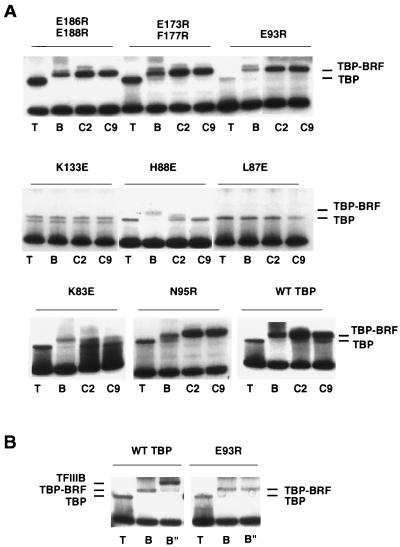
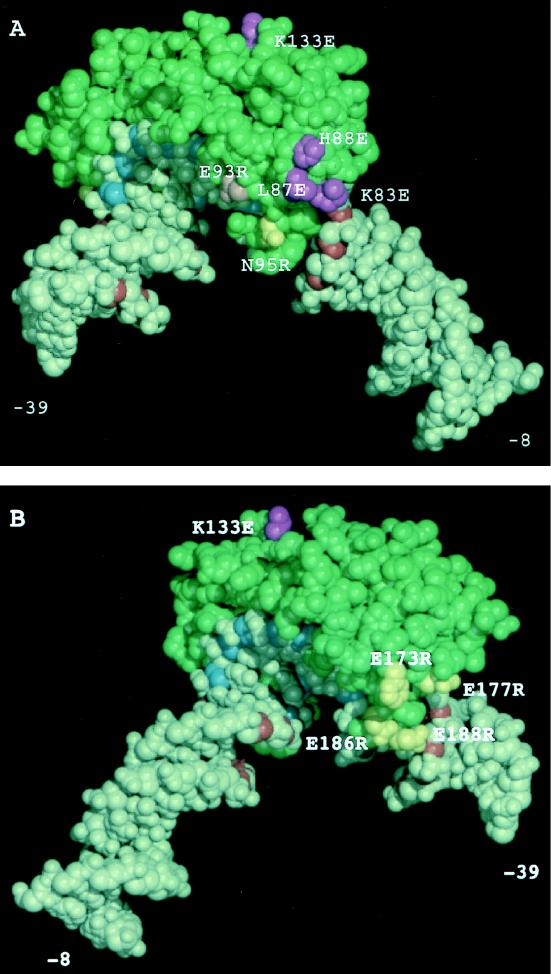
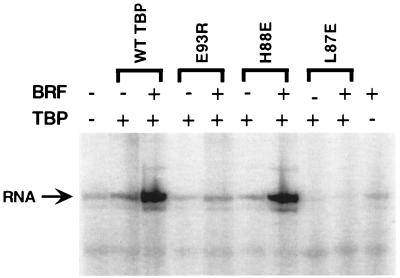
The RNA polymerase III factor TFIIIB forms a stable complex with DNA and can promote multiple rounds of initiation by polymerase. TFIIIB is composed of three subunits, the TATA binding protein (TBP), TFIIB-related factor (BRF), and B". Chemical footprinting, as well as mutagenesis of TBP, BRF, and promoter DNA, was used to probe the architecture of TFIIIB subunits bound to DNA. BRF bound to TBP-DNA through the nonconserved C-terminal region and required 15 bp downstream of the TATA box and as little as 1 bp upstream of the TATA box for stable complex formation. In contrast, formation of complete TFIIIB complexes required 15 bp both upstream and downstream of the TATA box. Hydroxyl radical footprinting of TFIIIB complexes and modeling the results to the TBP-DNA structure suggest that BRF and B" surround TBP on both faces of the TBP-DNA complex and provide an explanation for the exceptional stability of this complex. Competition for binding to TBP by BRF and either TFIIB or TFIIA suggests that BRF binds on the opposite face of the TBP-DNA complex from TFIIB and that the binding sites for TFIIA and BRF overlap. The positions of TBP mutations which are defective in binding BRF suggest that BRF binds to the top and N-terminal leg of TBP. One mutation on the N-terminal leg of TBP specifically affects the binding of the B" subunit.
Figures
References
-
- Bagby S, Kim S, Maldonado E, Tong K I, Reinberg D, Ikura M. Solution structure of the carboxy-terminal core domain of human TFIIB: similarity to cyclin A and interaction with TATA binding protein. Cell. 1995;82:857–867. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases