

Signaling through CD5 activates a pathway involving phosphatidylinositol 3-kinase, Vav, and Rac1 in human mature T lymphocytes
- PMID: 9488489
- PMCID: PMC108887
- DOI: 10.1128/MCB.18.3.1725
Signaling through CD5 activates a pathway involving phosphatidylinositol 3-kinase, Vav, and Rac1 in human mature T lymphocytes
Abstract
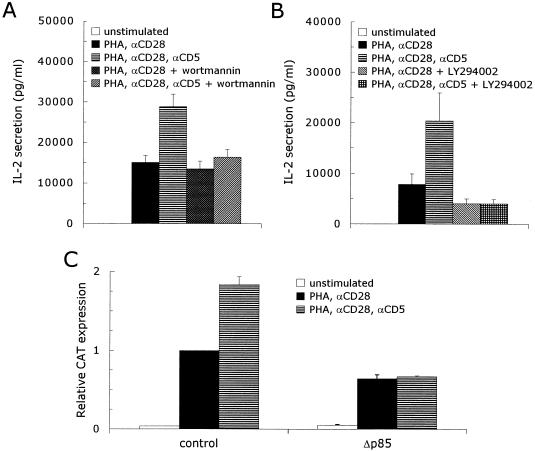
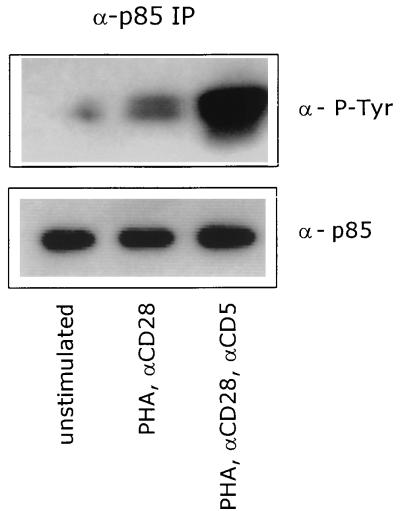
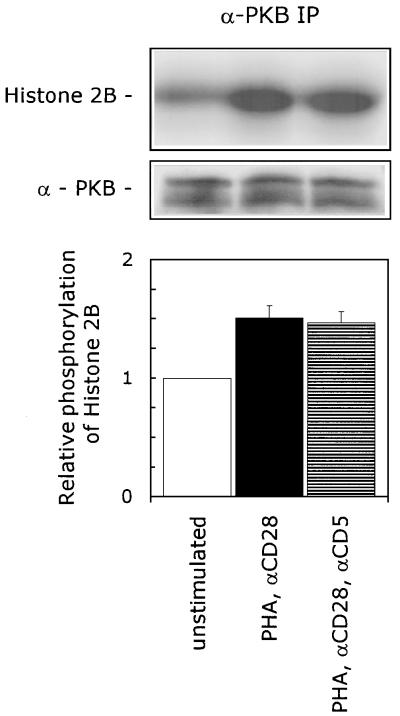
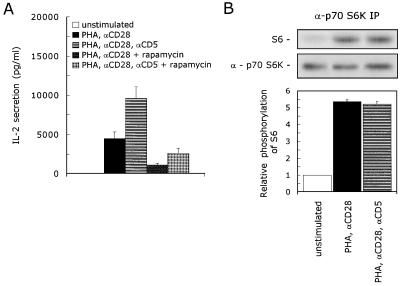
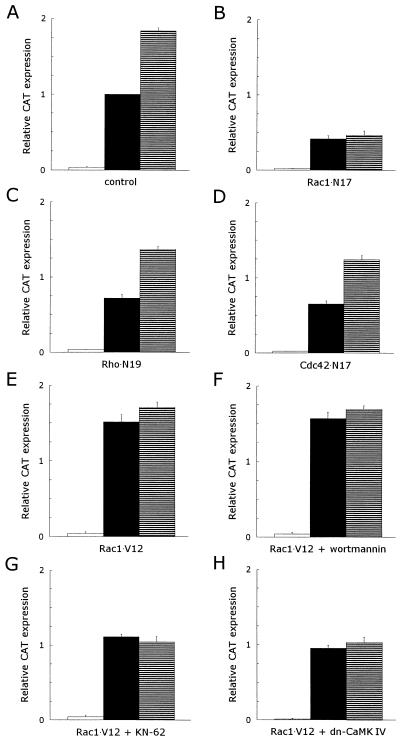
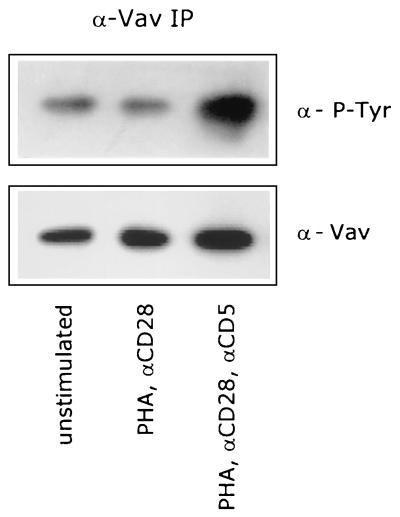
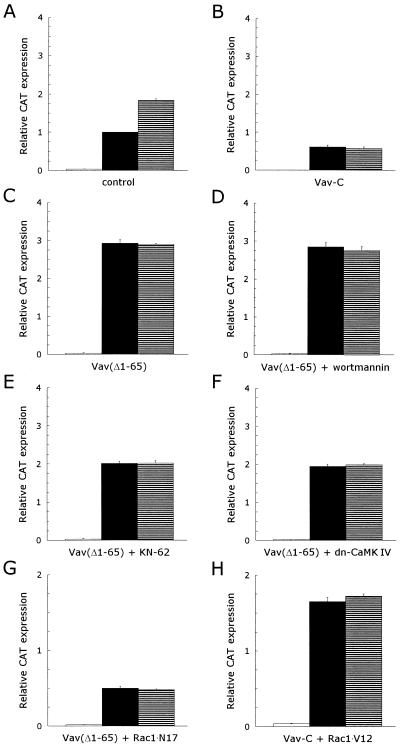
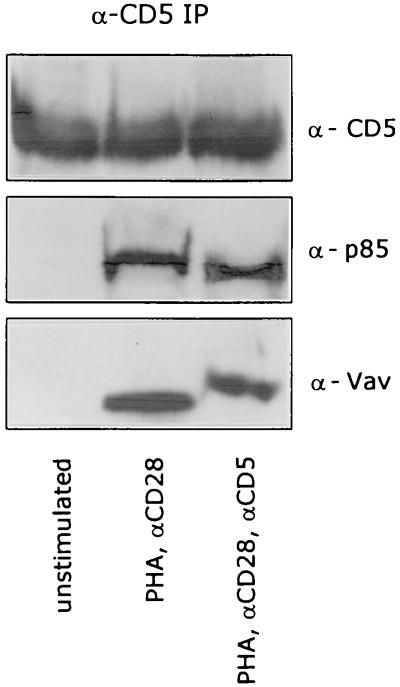
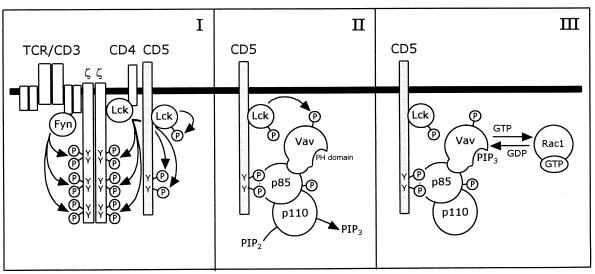
CD5 acts as a coreceptor on T lymphocytes and plays an important role in T-cell signaling and T-cell-B-cell interactions. Costimulation of T lymphocytes with anti-CD5 antibodies results in an increase of the intracellular Ca2+ levels, and subsequently in the activation of Ca2+/calmodulin-dependent (CaM) kinase type IV. In the present study, we have characterized the initial signaling pathway induced by anti-CD5 costimulation. The activation of phosphatidylinositol (PI) 3-kinase through tyrosine phosphorylation of its p85 subunit is a proximal event in the CD5-signaling pathway and leads to the activation of the lipid kinase activity of the p110 subunit. The PI 3-kinase inhibitors wortmannin and LY294002 inhibit the CD5-induced response as assessed in interleukin-2 (IL-2) secretion experiments. The expression of an inactivated Rac1 mutant (Rac1.N17) in T lymphocytes transfected with an IL-2 promoter-driven reporter construct also abrogates the response to CD5 costimulation, while the expression of a constitutively active Rac1 mutant (Rac1-V12) completely replaces the CD5 costimulatory signal. The Rac1-specific guanine nucleotide exchange factor Vav is heavily phosphorylated on tyrosine residues upon CD5 costimulation, which is a prerequisite for its activation. A role for Vav in the CD5-induced signaling pathway is further supported by the findings that the expression of a dominant negative Vav mutant (Vav-C) completely abolishes the response to CD5 costimulation while the expression of a constitutively active Vav mutant [Vav(delta1-65)] makes the CD5 costimulation signal superfluous. Wortmannin is unable to block the Vav(delta1-65)- or Rac1.V12-induced signals, indicating that both Vav and Rac1 function downstream from PI 3-kinase. Vav and Rac1 both act upstream from the CD5-induced activation of CaM kinase IV, since KN-62, an inhibitor of CaM kinases, and a dominant negative CaM kinase IV mutant block the Vav(delta1-65)-and Rac1.V12-mediated signals. We propose a model for the CD5-induced signaling pathway in which the PI 3-kinase lipid products, together with tyrosine phosphorylation, activate Vav, resulting in the activation of Rac1 by the Vav-mediated exchange of GDP for GTP.
Figures
References
-
- Alberola-Ila J, Places L, Cantrell D A, Vives J, Lozano F. Intracellular events involved in CD5-induced human T cell activation and proliferation. J Immunol. 1992;148:1287–1293. - PubMed
-
- al-Shami A, Bourgoin S G, Naccache P H. Granulocyte-macrophage colony-stimulating factor-activated signaling pathways in human neutrophils. I. Tyrosine phosphorylation-dependent stimulation of phosphatidylinositol 3-kinase and inhibition by phorbol esters. Blood. 1997;89:1035–1044. - PubMed
-
- Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous