

Differential targeting of vesicular stomatitis virus G protein and influenza virus hemagglutinin appears during myogenesis of L6 muscle cells
- PMID: 9490723
- PMCID: PMC2132697
- DOI: 10.1083/jcb.140.5.1101
Differential targeting of vesicular stomatitis virus G protein and influenza virus hemagglutinin appears during myogenesis of L6 muscle cells
Abstract
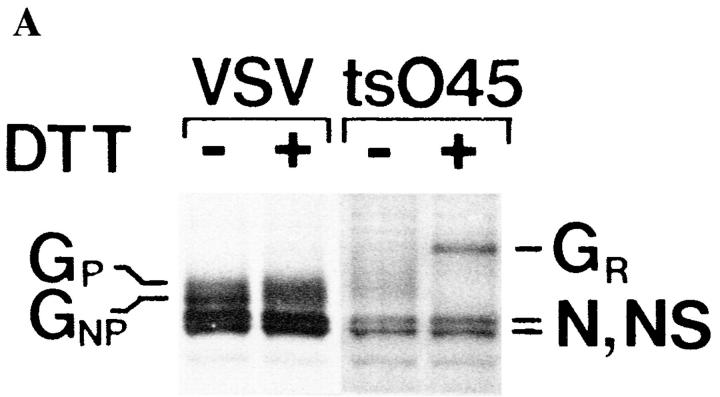
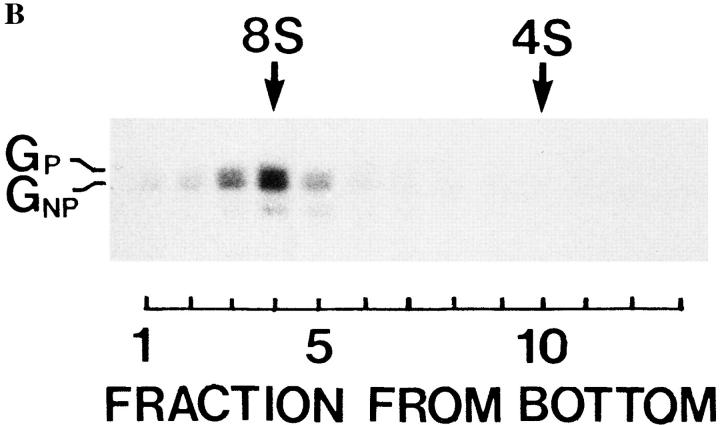
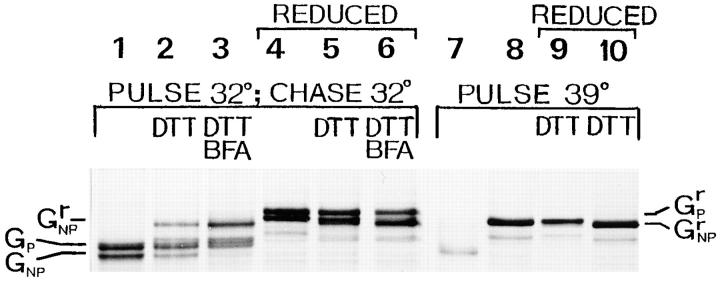
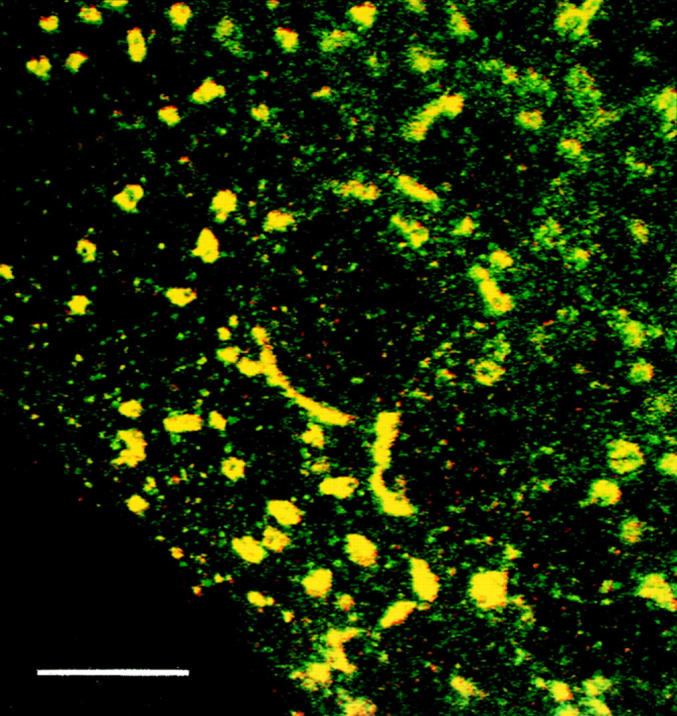
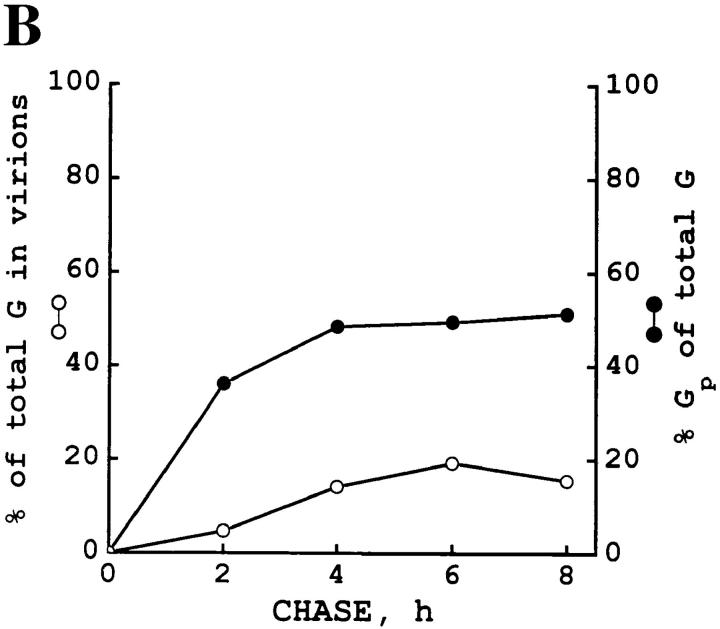
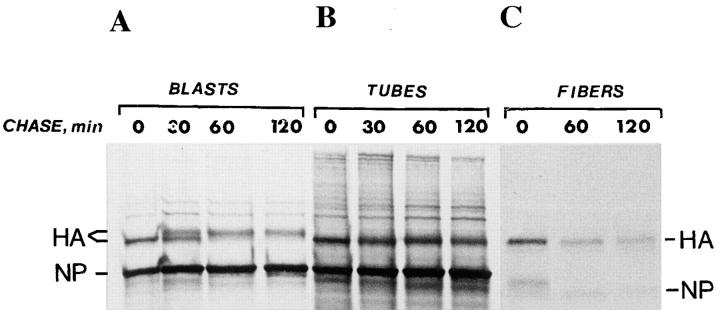
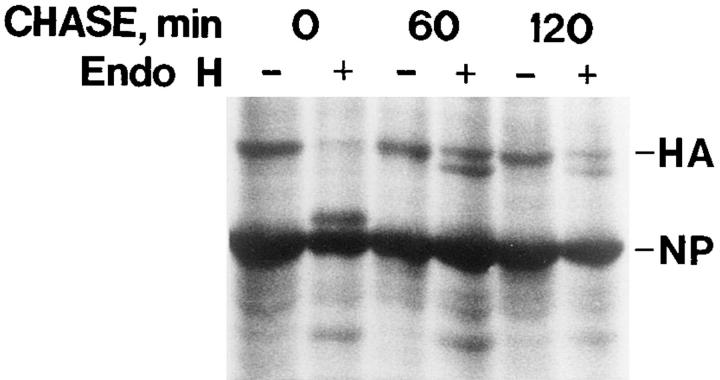
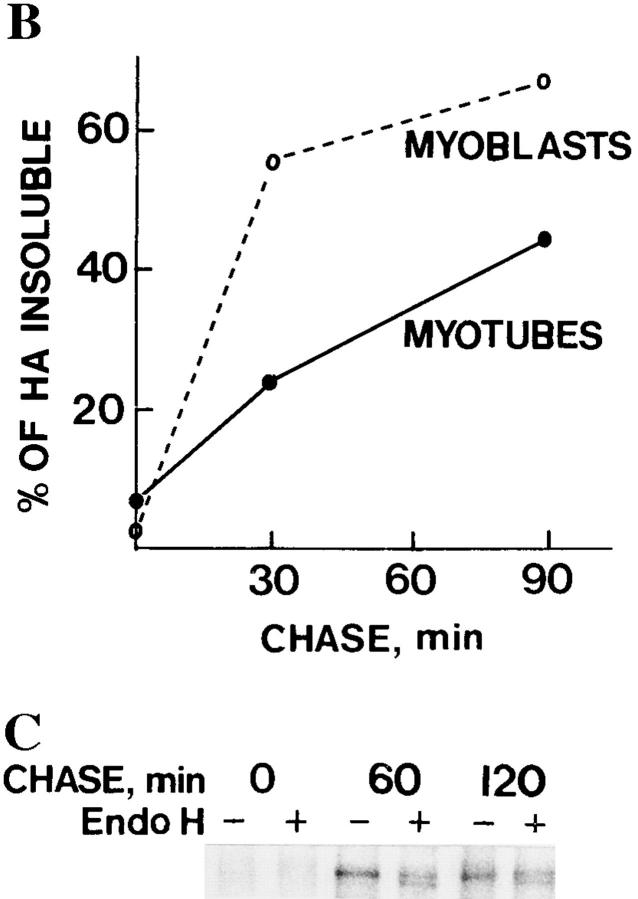
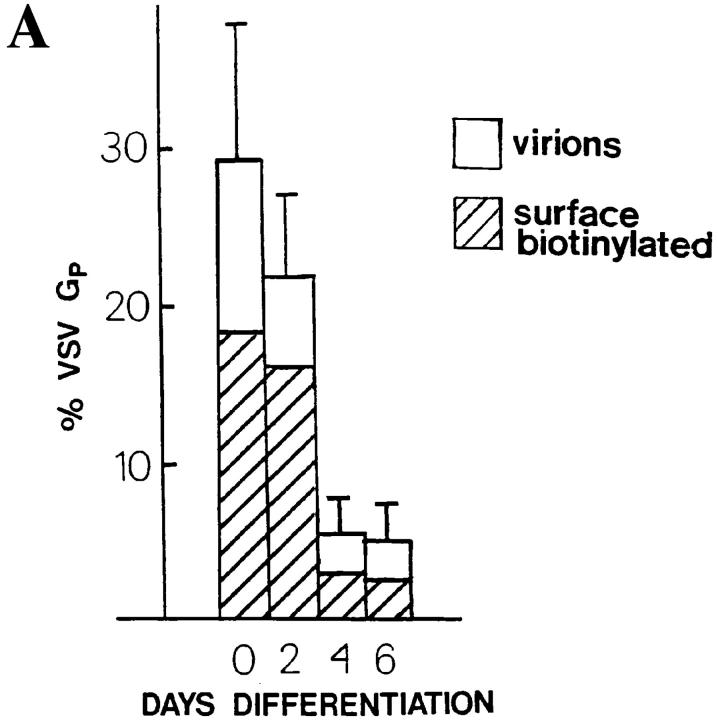
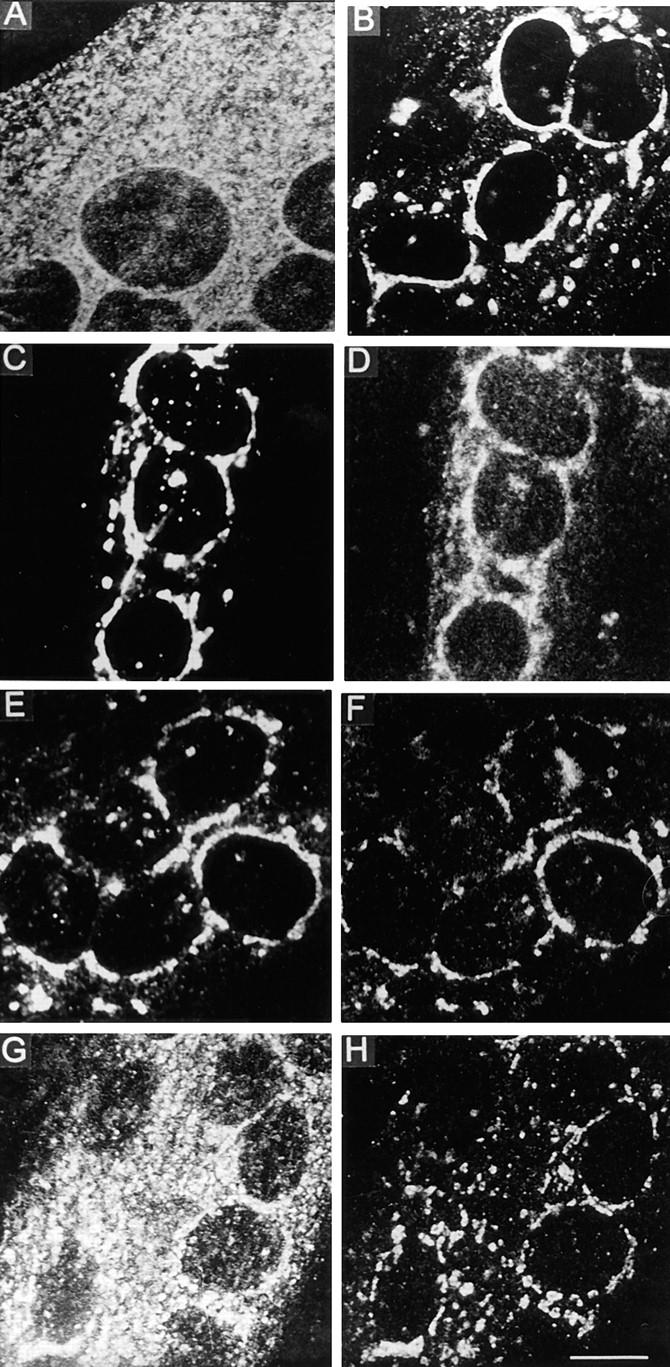
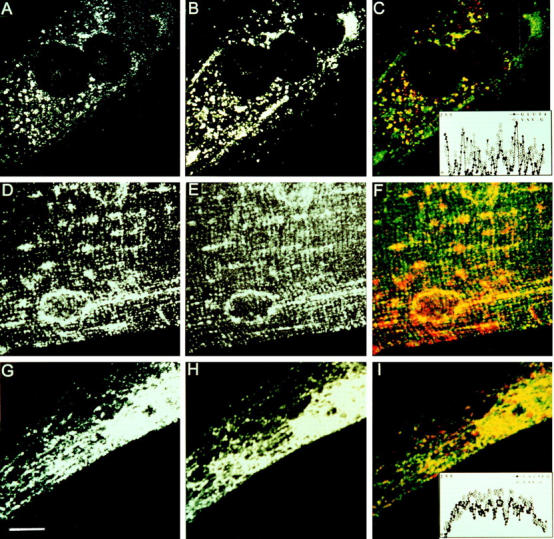
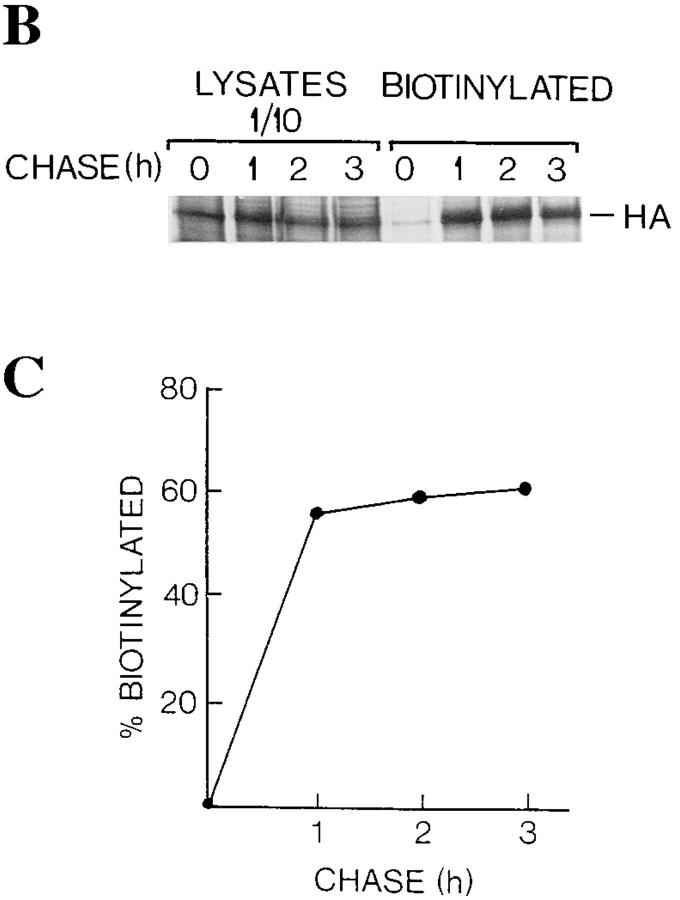
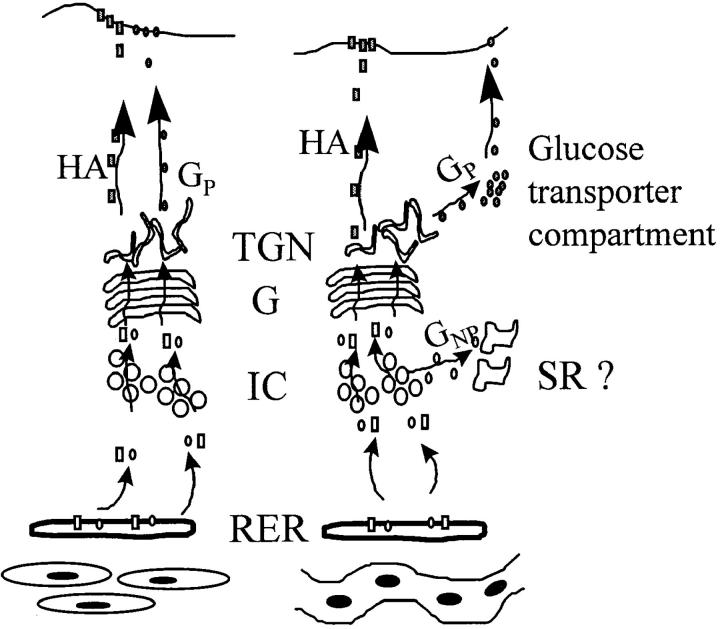
Exocytic organelles undergo profound reorganization during myoblast differentiation and fusion. Here, we analyzed whether glycoprotein processing and targeting changed during this process by using vesicular stomatitis virus (VSV) G protein and influenza virus hemagglutinin (HA) as models. After the induction of differentiation, the maturation and transport of the VSV G protein changed dramatically. Thus, only half of the G protein was processed and traveled through the Golgi, whereas the other half remained unprocessed. Experiments with the VSV tsO45 mutant indicated that the unprocessed form folded and trimerized normally and then exited the ER. It did not, however, travel through the Golgi since brefeldin A recalled it back to the ER. Influenza virus HA glycoprotein, on the contrary, acquired resistance to endoglycosidase H and insolubility in Triton X-100, indicating passage through the Golgi. Biochemical and morphological assays indicated that the HA appeared at the myotube surface. A major fraction of the Golgi-processed VSV G protein, however, did not appear at the myotube surface, but was found in intracellular vesicles that partially colocalized with the regulatable glucose transporter. Taken together, the results suggest that, during early myogenic differentiation, the VSV G protein was rerouted into developing, muscle-specific membrane compartments. Influenza virus HA, on the contrary, was targeted to the myotube surface.
Figures


Similar articles
-
Apical sorting of influenza hemagglutinin by transcytosis in retinal pigment epithelium.J Cell Sci. 1997 Aug;110 ( Pt 15):1717-27. doi: 10.1242/jcs.110.15.1717. J Cell Sci. 1997. PMID: 9264459
-
Retargeting of viral glycoproteins into a non-exporting compartment during the myogenic differentiation of rat L6 cells.Cell Tissue Res. 2002 Jun;308(3):381-90. doi: 10.1007/s00441-002-0556-5. Epub 2002 Apr 25. Cell Tissue Res. 2002. PMID: 12107431
-
Morphological analysis of protein transport from the ER to Golgi membranes in digitonin-permeabilized cells: role of the P58 containing compartment.J Cell Biol. 1992 Dec;119(5):1097-116. doi: 10.1083/jcb.119.5.1097. J Cell Biol. 1992. PMID: 1447290 Free PMC article.
-
Analysis of protein transport through the Golgi in a reconstituted cell-free system.J Electron Microsc Tech. 1991 Feb;17(2):150-64. doi: 10.1002/jemt.1060170204. J Electron Microsc Tech. 1991. PMID: 1901603 Review.
-
Viral membrane fusion.Nat Struct Mol Biol. 2008 Jul;15(7):690-8. doi: 10.1038/nsmb.1456. Nat Struct Mol Biol. 2008. PMID: 18596815 Free PMC article. Review.
Cited by
-
Analysis of GLUT4 distribution in whole skeletal muscle fibers: identification of distinct storage compartments that are recruited by insulin and muscle contractions.J Cell Biol. 1998 Sep 21;142(6):1429-46. doi: 10.1083/jcb.142.6.1429. J Cell Biol. 1998. PMID: 9744875 Free PMC article.
-
Insulin resistance and the disruption of Glut4 trafficking in skeletal muscle.J Clin Invest. 2001 May;107(10):1211-3. doi: 10.1172/JCI13020. J Clin Invest. 2001. PMID: 11375407 Free PMC article. No abstract available.
-
Vesicle budding from endoplasmic reticulum is involved in calsequestrin routing to sarcoplasmic reticulum of skeletal muscles.Biochem J. 2004 Apr 15;379(Pt 2):505-12. doi: 10.1042/BJ20031875. Biochem J. 2004. PMID: 14728599 Free PMC article.
-
Golgi complex reorganization during muscle differentiation: visualization in living cells and mechanism.Mol Biol Cell. 2001 Apr;12(4):795-808. doi: 10.1091/mbc.12.4.795. Mol Biol Cell. 2001. PMID: 11294887 Free PMC article.
-
Loss of a Clueless-dGRASP complex results in ER stress and blocks Integrin exit from the perinuclear endoplasmic reticulum in Drosophila larval muscle.Biol Open. 2015 Apr 10;4(5):636-48. doi: 10.1242/bio.201511551. Biol Open. 2015. PMID: 25862246 Free PMC article.
References
-
- Balch WE, Keller DS. ATP-coupled transport of vesicular stomatitis virus G protein. Functional boundaries of secretory compartments. J Biol Chem. 1986;261:14690–14696. - PubMed
-
- Baron MD, Garoff H. Mannosidase II and the 135 kDa Golgi-specific antigen recognized monoclonal antibody 53FC3 are the same dimeric protein. J Biol Chem. 1990;265:19928–19931. - PubMed
