

Moderately High Temperatures Inhibit Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase (Rubisco) Activase-Mediated Activation of Rubisco
- PMID: 9490757
- PMCID: PMC35111
- DOI: 10.1104/pp.116.2.539
Moderately High Temperatures Inhibit Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase (Rubisco) Activase-Mediated Activation of Rubisco
Abstract
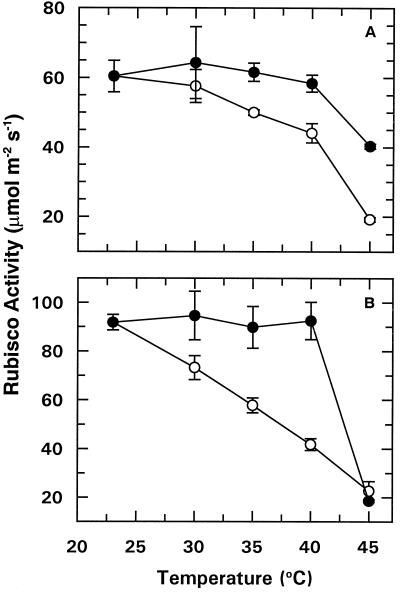
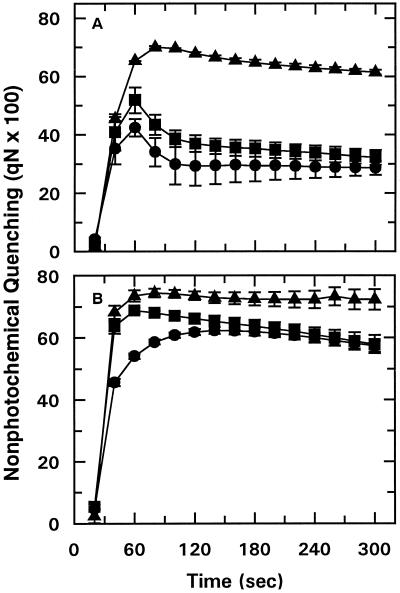
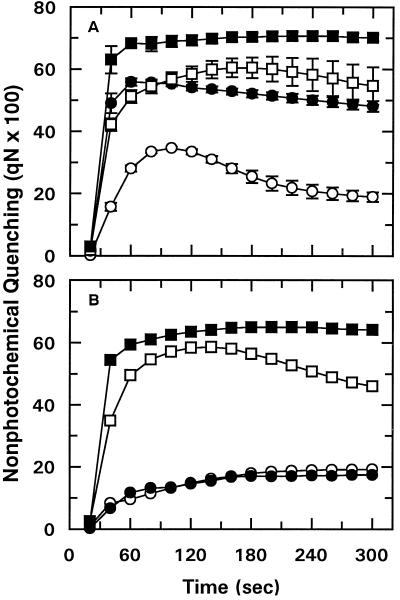
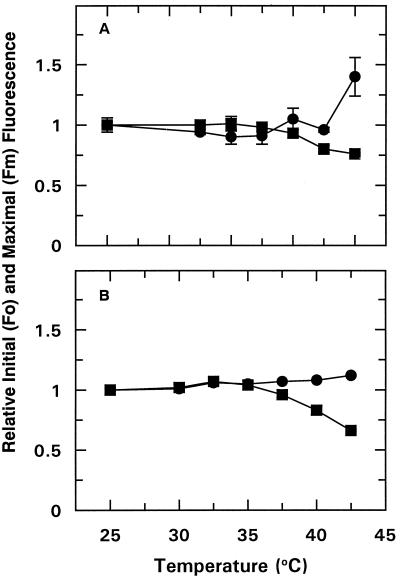
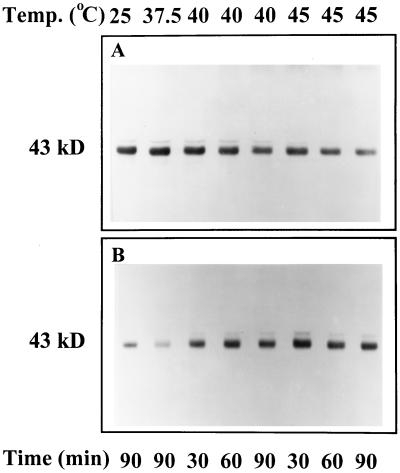
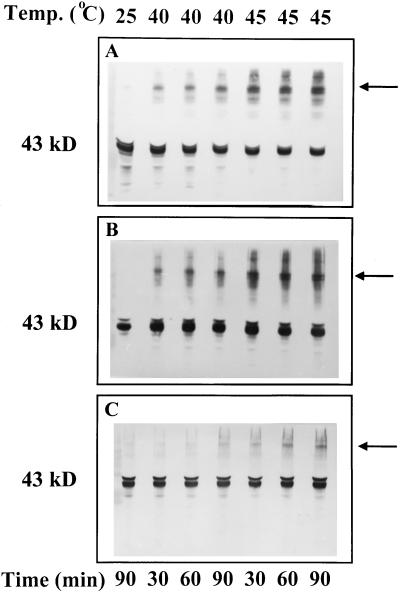
We tested the hypothesis that light activation of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) is inhibited by moderately elevated temperature through an effect on Rubisco activase. When cotton (Gossypium hirsutum L.) or wheat (Triticum aestivum L.) leaf tissue was exposed to increasing temperatures in the light, activation of Rubisco was inhibited above 35 and 30 degreesC, respectively, and the relative inhibition was greater for wheat than for cotton. The temperature-induced inhibition of Rubisco activation was fully reversible at temperatures below 40 degreesC. In contrast to activation state, total Rubisco activity was not affected by temperatures as high as 45 degreesC. Nonphotochemical fluorescence quenching increased at temperatures that inhibited Rubisco activation, consistent with inhibition of Calvin cycle activity. Initial and maximal chlorophyll fluorescence were not significantly altered until temperatures exceeded 40 degreesC. Thus, electron transport, as measured by Chl fluorescence, appeared to be more stable to moderately elevated temperatures than Rubisco activation. Western-blot analysis revealed the formation of high-molecular-weight aggregates of activase at temperatures above 40 degreesC for both wheat and cotton when inhibition of Rubisco activation was irreversible. Physical perturbation of other soluble stromal enzymes, including Rubisco, phosphoribulokinase, and glutamine synthetase, was not detected at the elevated temperatures. Our evidence indicates that moderately elevated temperatures inhibit light activation of Rubisco via a direct effect on Rubisco activase.
Figures

Similar articles
-
Inhibition and acclimation of photosynthesis to heat stress is closely correlated with activation of ribulose-1,5-bisphosphate Carboxylase/Oxygenase.Plant Physiol. 1999 May;120(1):173-82. doi: 10.1104/pp.120.1.173. Plant Physiol. 1999. PMID: 10318695 Free PMC article.
-
Effect of heat stress on the inhibition and recovery of the ribulose-1,5-bisphosphate carboxylase/oxygenase activation state.Planta. 2000 Dec;212(1):67-74. doi: 10.1007/s004250000364. Planta. 2000. PMID: 11219585
-
Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2.Proc Natl Acad Sci U S A. 2000 Nov 21;97(24):13430-5. doi: 10.1073/pnas.230451497. Proc Natl Acad Sci U S A. 2000. PMID: 11069297 Free PMC article.
-
The temperature response of C(3) and C(4) photosynthesis.Plant Cell Environ. 2007 Sep;30(9):1086-106. doi: 10.1111/j.1365-3040.2007.01682.x. Plant Cell Environ. 2007. PMID: 17661749 Review.
-
Rubisco, Rubisco activase, and global climate change.J Exp Bot. 2008;59(7):1581-95. doi: 10.1093/jxb/ern053. Epub 2008 Apr 23. J Exp Bot. 2008. PMID: 18436544 Review.
Cited by
-
Characterization of common and distinctive adjustments of wild barley leaf proteome under drought acclimation, heat stress and their combination.Plant Mol Biol. 2015 Mar;87(4-5):459-71. doi: 10.1007/s11103-015-0291-4. Epub 2015 Feb 3. Plant Mol Biol. 2015. PMID: 25647426
-
A Conserved Sequence from Heat-Adapted Species Improves Rubisco Activase Thermostability in Wheat.Plant Physiol. 2019 Sep;181(1):43-54. doi: 10.1104/pp.19.00425. Epub 2019 Jun 12. Plant Physiol. 2019. PMID: 31189658 Free PMC article.
-
Inhibition and acclimation of photosynthesis to heat stress is closely correlated with activation of ribulose-1,5-bisphosphate Carboxylase/Oxygenase.Plant Physiol. 1999 May;120(1):173-82. doi: 10.1104/pp.120.1.173. Plant Physiol. 1999. PMID: 10318695 Free PMC article.
-
The different patterns of post-heat stress responses in wheat genotypes: the role of the transthylakoid proton gradient in efficient recovery of leaf photosynthetic capacity.Photosynth Res. 2021 Dec;150(1-3):179-193. doi: 10.1007/s11120-020-00812-0. Epub 2021 Jan 3. Photosynth Res. 2021. PMID: 33393064
-
Tandemly duplicated Rubisco activase genes of cereals show differential evolution and response to heat stress.Plant Mol Biol. 2024 Dec 21;115(1):10. doi: 10.1007/s11103-024-01515-z. Plant Mol Biol. 2024. PMID: 39708272
References
-
- Andrews TJ, Hudson GS, Mate CJ, von Caemmerer S, Evans JR, Avridsson YBC. Rubisco, consequences of altering its expression and activation in transgenic plants. J Exp Bot. 1995;46:1293–1300.
-
- Berry JA, Björkman O. Photosynthetic response and adaptation to temperature in higher plants. Annu Rev Plant Physiol. 1980;31:491–543.
-
- Bilger W, Schreiber U, Lange OL. Chlorophyll fluorescence as an indicator of heat induced limitation of photosynthesis in Arbutus unedo L. In: Tenhunen JD, Catarino FM, Lange OL, editors. Plant Response to Stress. Berlin: Springer; 1987. pp. 391–399.
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of proteins utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–259. - PubMed
LinkOut - more resources
Full Text Sources
