

The open reading frame (ORF) 50a gene product regulates ORF 57 gene expression in herpesvirus saimiri
- PMID: 9499050
- PMCID: PMC109489
- DOI: 10.1128/JVI.72.3.1967-1973.1998
The open reading frame (ORF) 50a gene product regulates ORF 57 gene expression in herpesvirus saimiri
Abstract
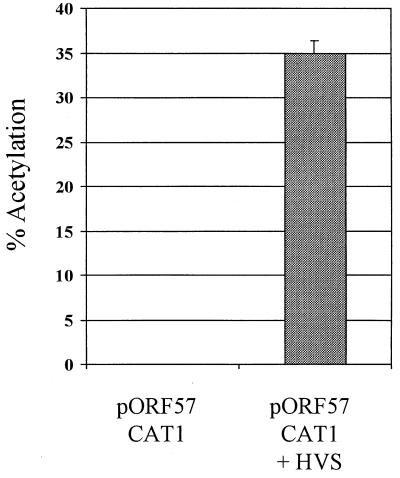
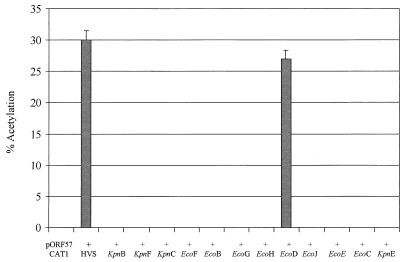
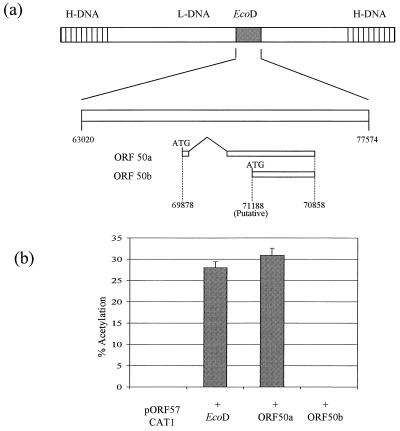
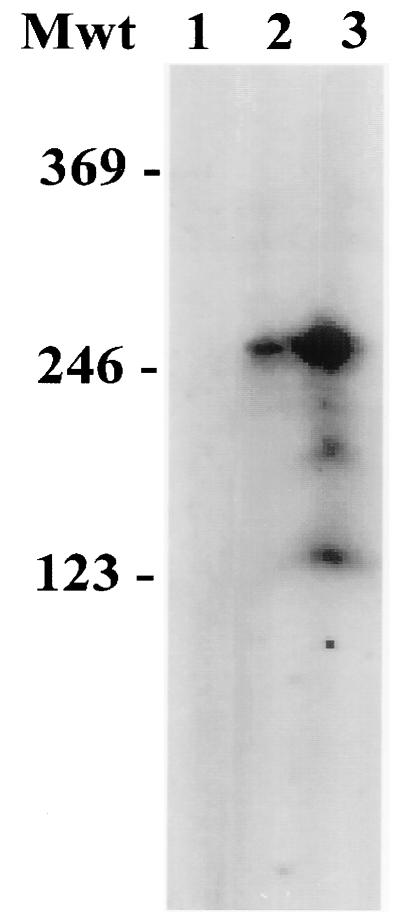
We have previously demonstrated that open reading frame (ORF) 50 and ORF 57 encode transcriptional regulating genes in herpesvirus saimiri. ORF 50, a homolog of Epstein-Barr virus R protein, is a sequence-specific transactivator, whereas ORF 57 acts posttranscriptionally. In this report, we demonstrate that the ORF 57 gene is regulated by the ORF 50a gene product. We show that the ORF 57 gene is expressed at basal levels early in the virus replication cycle and that thereafter it is transactivated by the ORF 50a gene product, due to an increase in RNA levels. As it has been shown that the ORF 57 gene product downregulates ORF 50a due to the presence of its intron, these combined observations identify a feedback mechanism modulating gene expression in herpesvirus saimiri, whereby ORF 50a transcription is downregulated by the ORF 57 gene product, a gene which it specifically transactivates. Furthermore, we propose that the intron-containing ORF 57 gene downregulates itself by the same mechanism as that for ORF 50a, as both genes are downregulated at similar times during the replication cycle.
Figures

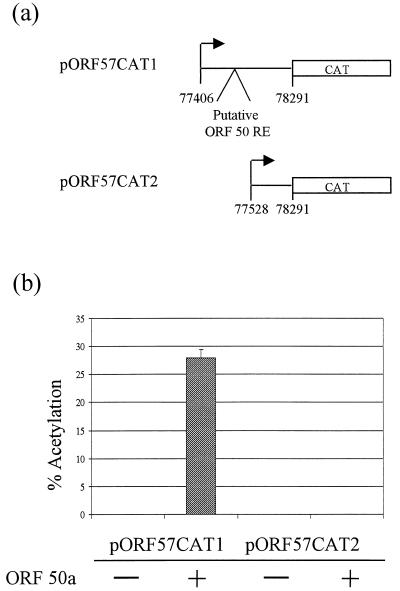
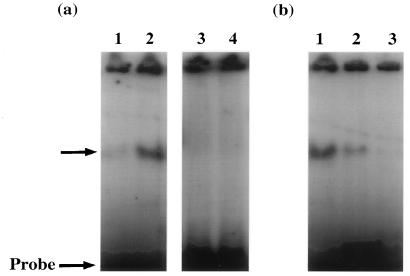
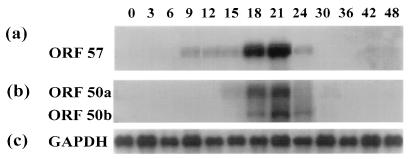

Similar articles
-
The Herpesvirus saimiri replication and transcription activator acts synergistically with CCAAT enhancer binding protein alpha to activate the DNA polymerase promoter.J Virol. 2005 Nov;79(21):13548-60. doi: 10.1128/JVI.79.21.13548-13560.2005. J Virol. 2005. PMID: 16227275 Free PMC article.
-
The immediate-early gene product encoded by open reading frame 57 of herpesvirus saimiri modulates gene expression at a posttranscriptional level.J Virol. 1998 Jan;72(1):857-61. doi: 10.1128/JVI.72.1.857-861.1998. J Virol. 1998. PMID: 9420300 Free PMC article.
-
The herpesvirus saimiri Rta gene autostimulates via binding to a non-consensus response element.J Gen Virol. 2005 Mar;86(Pt 3):581-587. doi: 10.1099/vir.0.80723-0. J Gen Virol. 2005. PMID: 15722517
-
Bovine herpesvirus 4: genomic organization and relationship with two other gammaherpesviruses, Epstein-Barr virus and herpesvirus saimiri.Vet Microbiol. 1996 Nov;53(1-2):79-89. doi: 10.1016/s0378-1135(96)01236-9. Vet Microbiol. 1996. PMID: 9011000 Review.
-
gamma-2 Herpes virus post-transcriptional gene regulation.Clin Microbiol Infect. 2006 Feb;12(2):110-7. doi: 10.1111/j.1469-0691.2005.01317.x. Clin Microbiol Infect. 2006. PMID: 16441447 Review.
Cited by
-
Open reading frame 50 protein of Kaposi's sarcoma-associated herpesvirus directly activates the viral PAN and K12 genes by binding to related response elements.J Virol. 2002 Apr;76(7):3168-78. doi: 10.1128/jvi.76.7.3168-3178.2002. J Virol. 2002. PMID: 11884541 Free PMC article.
-
Herpesvirus saimiri.Philos Trans R Soc Lond B Biol Sci. 2001 Apr 29;356(1408):545-67. doi: 10.1098/rstb.2000.0780. Philos Trans R Soc Lond B Biol Sci. 2001. PMID: 11313011 Free PMC article. Review.
-
The Herpesvirus saimiri replication and transcription activator acts synergistically with CCAAT enhancer binding protein alpha to activate the DNA polymerase promoter.J Virol. 2005 Nov;79(21):13548-60. doi: 10.1128/JVI.79.21.13548-13560.2005. J Virol. 2005. PMID: 16227275 Free PMC article.
-
The activation domain of herpesvirus saimiri R protein interacts with the TATA-binding protein.J Virol. 1999 Dec;73(12):9756-63. doi: 10.1128/JVI.73.12.9756-9763.1999. J Virol. 1999. PMID: 10559285 Free PMC article.
-
Control of Rta expression critically determines transcription of viral and cellular genes following gammaherpesvirus infection.J Gen Virol. 2007 Jun;88(Pt 6):1689-1697. doi: 10.1099/vir.0.82548-0. J Gen Virol. 2007. PMID: 17485528 Free PMC article.
References
-
- Albrecht J C, Fleckenstein B. Structural organization of the conserved gene block of herpesvirus saimiri coding for DNA polymerase, glycoprotein B, and major DNA binding protein. Virology. 1990;174:533–542. - PubMed
-
- Bublot M, Manet E, Lequarre A S, Albrecht J C, Nicholas J, Fleckenstein B, Pastoret P P, Thiry E. Genetic relationships between bovine herpesvirus 4 and the gamma-herpesviruses Epstein-Barr and herpesvirus saimiri. Virology. 1992;190:654–665. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
