

Chimeric measles viruses with a foreign envelope
- PMID: 9499071
- PMCID: PMC109510
- DOI: 10.1128/JVI.72.3.2150-2159.1998
Chimeric measles viruses with a foreign envelope
Abstract
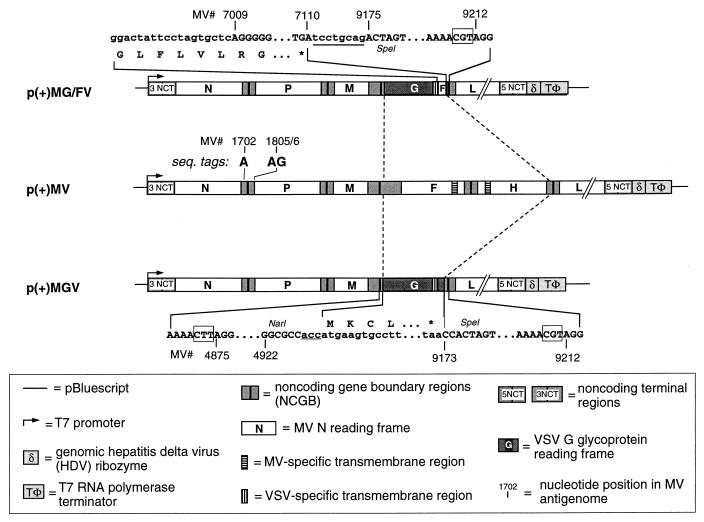
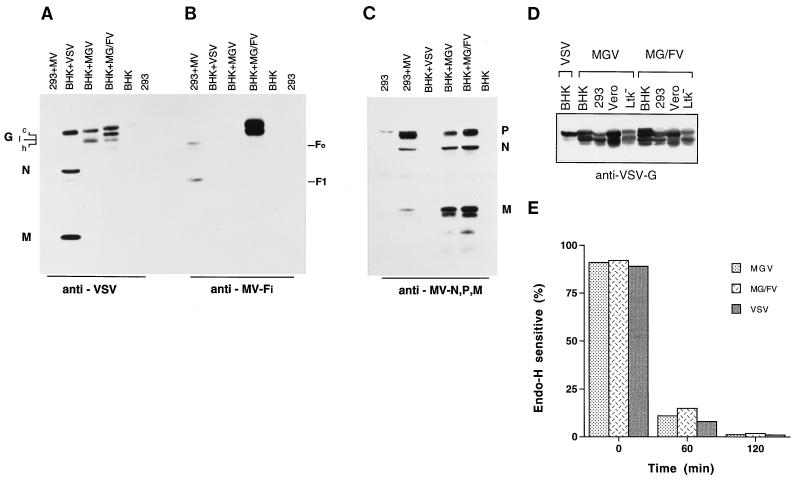
Measles virus (MV) and vesicular stomatitis virus (VSV) are both members of the Mononegavirales but are only distantly related. We generated two genetically stable chimeric viruses. In MGV, the reading frames of the MV envelope glycoproteins H and F were substituted by a single reading frame encoding the VSV G glycoprotein; MG/FV is similar but encodes a G/F hybrid in which the VSV G cytoplasmic tail was replaced by that of MV F. In contrast to MG/FV, MGV virions do not contain the MV matrix (M) protein. This demonstrates that virus assembly is possible in the absence of M; conversely, the cytoplasmic domain of F allows incorporation of M and enhances assembly. The formation of chimeric viruses was substantially delayed and the titers obtained were reduced about 50-fold in comparison to standard MV. In the novel chimeras, transcription and replication are mediated by the MV ribonucleoproteins but the envelope glycoproteins dictate the host range. Mice immunized with the chimeric viruses were protected against lethal doses of wild-type VSV. These findings suggest that it is feasible to construct MV variants bearing a variety of different envelopes for use as vaccines or for gene therapeutic purposes.
Figures
References
-
- Billeter M A, Cattaneo R, Schmid A, Eschle D, Kaelin K, Rebmann G, Udem S A, Sheppard R D, Baczko K, Liebert U G, Schneider-Schaulies S, Brinckmann U, ter Meulen V. Host and viral features in persistent measles virus infections of the brain. In: Kolakofsky D, Mahy B W J, editors. Genetics and pathogenicity of negative strand viruses. Amsterdam, The Netherlands: Elsevier Science Publishers BV; 1989. pp. 356–366.
-
- Burge B W, Pfefferkorn E R. Phenotypic mixing between group A arboviruses. Nature. 1966;210:1397–1399. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
