

A 2.9-kilobase noncoding nuclear RNA functions in the establishment of persistent Hz-1 viral infection
- PMID: 9499081
- PMCID: PMC109520
- DOI: 10.1128/JVI.72.3.2233-2245.1998
A 2.9-kilobase noncoding nuclear RNA functions in the establishment of persistent Hz-1 viral infection
Abstract
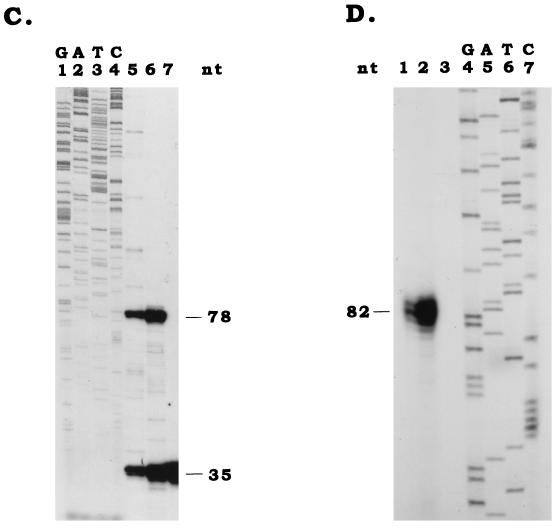
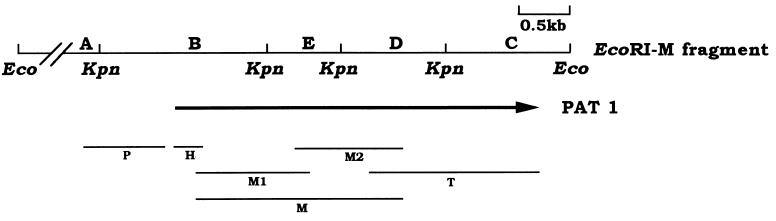
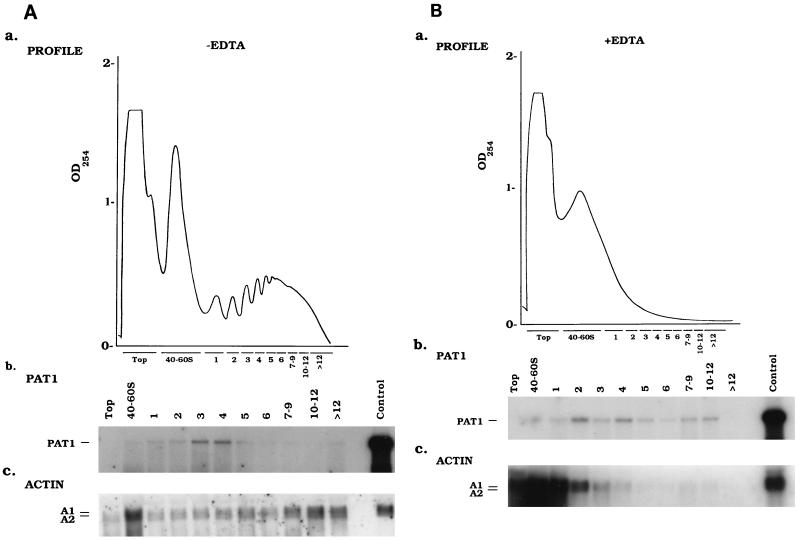
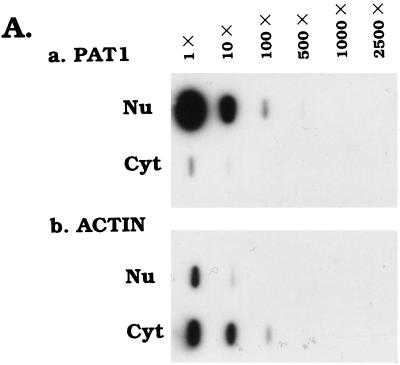
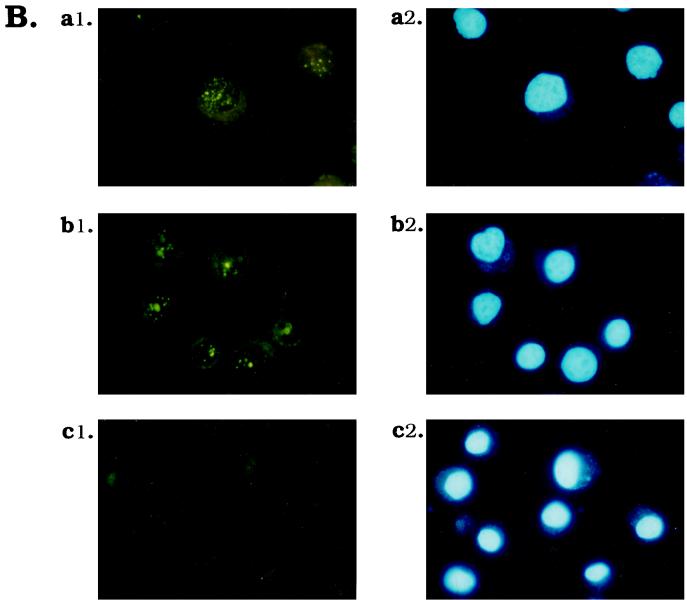
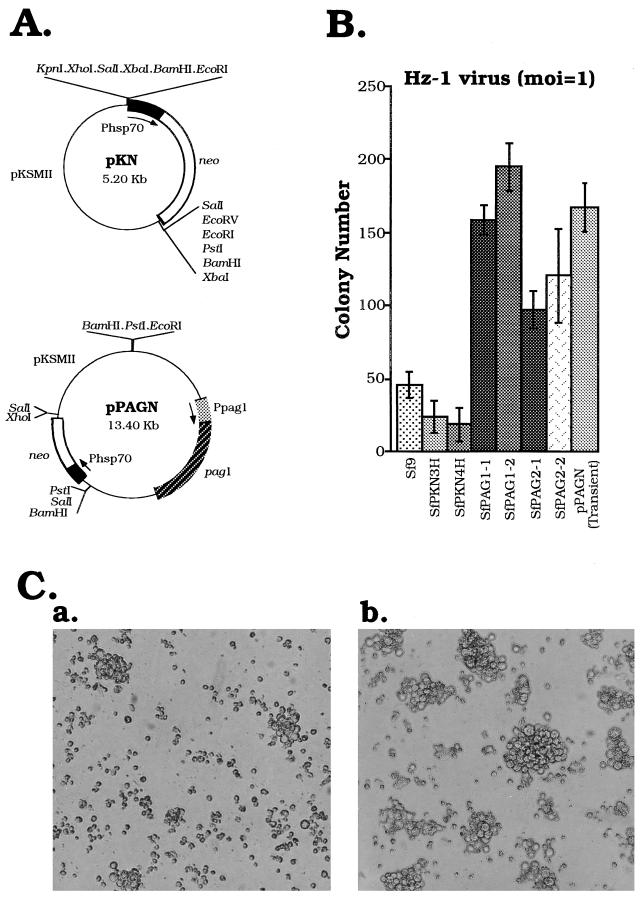
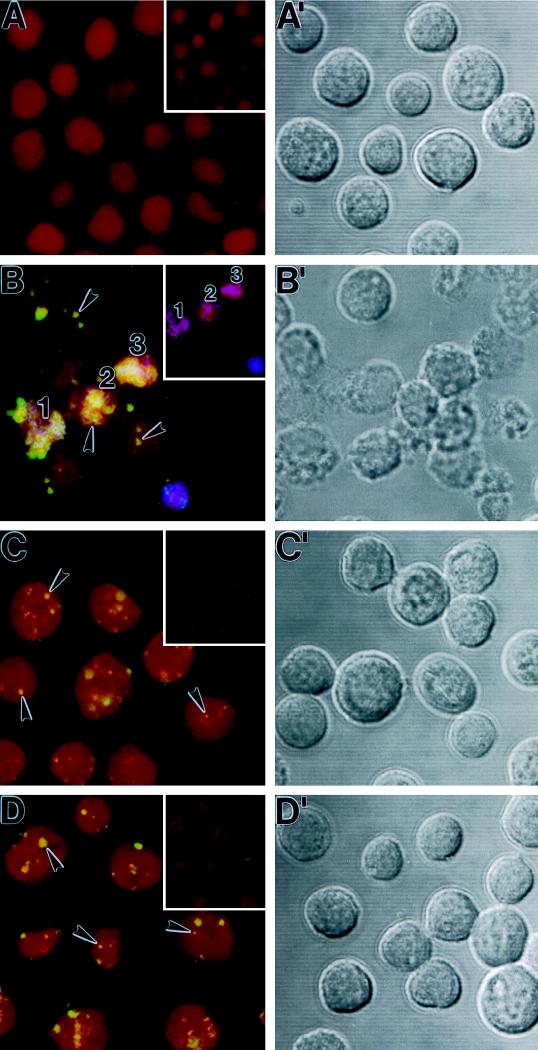
Differential viral gene expression during both productive and persistent infections of Hz-1 virus in insect cells was elucidated. Despite more than 100 viral transcripts being expressed during productive viral infection, massive viral gene shutoff was observed during viral persistency, leaving the 2.9-kb persistence-associated transcript 1 (PAT1) as the only detectable viral RNA. Persistence-associated gene 1 (pag1), which encodes PAT1, was cloned and found to contain no significant open reading frames. PAT1 is not associated with the cellular translation machinery and is located exclusively in the nucleus. Further experiments showed that PAT1 is functional in the establishment of persistent Hz-1 viral infection in the cells. All the evidence collectively indicates that PAT1 is a novel nuclear transcript of viral origin. Our results showed that although PAT1 and XIST RNA, a mammalian X-inactive specific transcript, are transcribed by different genes, they have interesting similarities.
Figures



References
-
- Blissard G W, Kogan P H, Wei R, Rohrmann G F. A synthetic early promoter from a baculovirus: roles of the TATA box and conserved start site CAGT sequence in basal levels of transcription. Virology. 1992;190:783–793. - PubMed
-
- Blissard G W, Rohrmann G F. Baculovirus diversity and molecular biology. Annu Rev Entomol. 1990;35:127–155. - PubMed
-
- Brockdorff N, Ashworth A, Graham F K, McCabe V M, Norris D P, Cooper P J, Swift S, Rastan S. The product of the mouse XIST gene is a 15 kb inactive X specific transcript containing no conserved ORF and located in the nucleus. Cell. 1992;71:515–526. - PubMed
-
- Brown C J, Hendrich B D, Rupert J L, Lafreniere R G, Xing Y, Lawrence J, Willard H F. The human XIST gene: analysis of a 17 kb inactive X-specific RNA that contains conserved repeats and is highly localized within the nucleus. Cell. 1992;71:527–542. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
