

Distinct pathways for tumor necrosis factor alpha and ceramides in human cytomegalovirus infection
- PMID: 9499092
- PMCID: PMC109531
- DOI: 10.1128/JVI.72.3.2316-2322.1998
Distinct pathways for tumor necrosis factor alpha and ceramides in human cytomegalovirus infection
Abstract
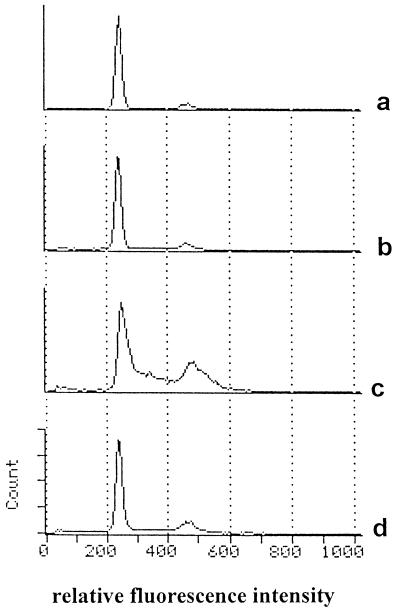
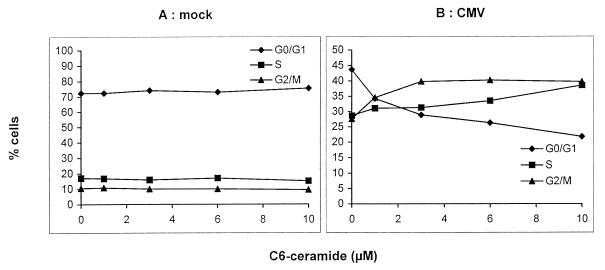
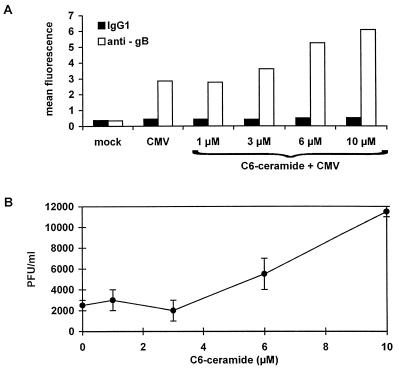
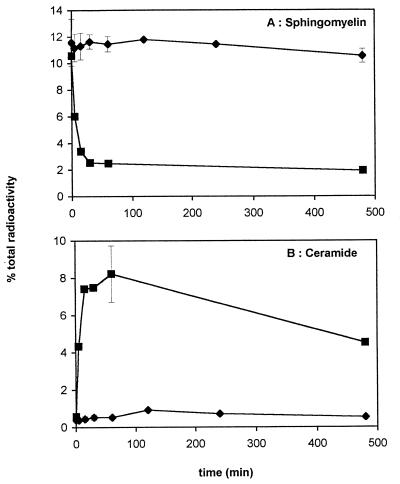
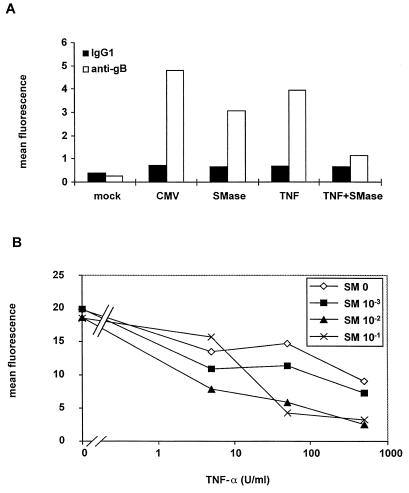
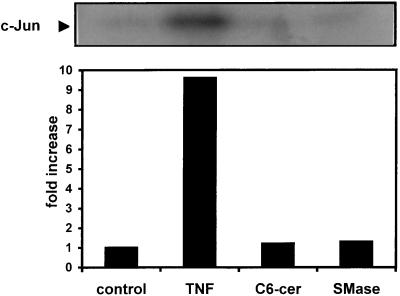
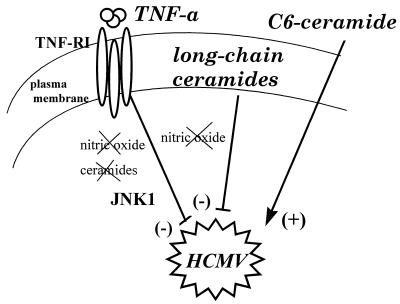
Human cytomegalovirus (HCMV) infection can be fatal to immunocompromised individuals. We have previously reported that gamma interferon and tumor necrosis factor alpha (TNF-alpha) synergistically inhibit HCMV replication in vitro. Ceramides have been described as second messengers induced by TNF-alpha. To investigate the mechanisms involved in the inhibition of HCMV by TNF-alpha, in the present study we have analyzed ceramide production by U373 MG astrocytoma cells and the effects of TNF-alpha versus ceramides on HCMV replication. Our results show that U373 MG cells did not produce ceramides upon incubation with TNF-alpha. Moreover, long-chain ceramides induced by treatment with exogenous bacterial sphingomyelinase inhibited HCMV replication in synergy with TNF-alpha. Surprisingly, short-chain permeant C6-ceramide increased viral replication. Our results show that the anti-HCMV activity of TNF-alpha is independent of ceramides. In addition, our results suggest that TNF-alpha and endogenous long-chain ceramides use separate pathways of cell signalling to inhibit HCMV replication, while permeant C6-ceramide appears to activate a third pathway leading to an opposite effect.
Figures
References
-
- Bligh E G, Dyer W J. A rapid method of total lipid extraction and purification. Can J Biochem Physiol. 1959;37:911–918. - PubMed
-
- Bresnahan W A, Boldogh I, Aubrey Thomson E, Albrecht T. Human cytomegalovirus inhibits cellular DNA synthesis and arrests productively infected cells in late G1. Virology. 1996;224:150–160. - PubMed
-
- Britt W J, Alford C A. Cytomegalovirus. In: Fields B N, editor. Virology. 3rd ed. Philadelphia, Pa: Lippincott-Raven Publishers; 1996. pp. 2493–2523.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
