

Extended minus-strand DNA as template for R-U5-mediated second-strand transfer in recombinational rescue of primer binding site-modified retroviral vectors
- PMID: 9499117
- PMCID: PMC109556
- DOI: 10.1128/JVI.72.3.2519-2525.1998
Extended minus-strand DNA as template for R-U5-mediated second-strand transfer in recombinational rescue of primer binding site-modified retroviral vectors
Abstract
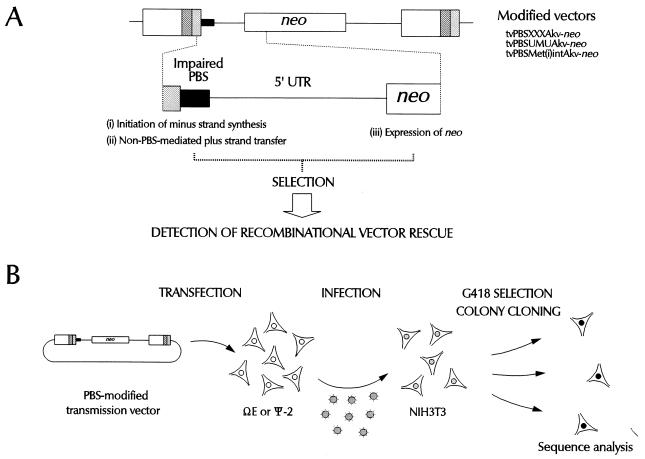
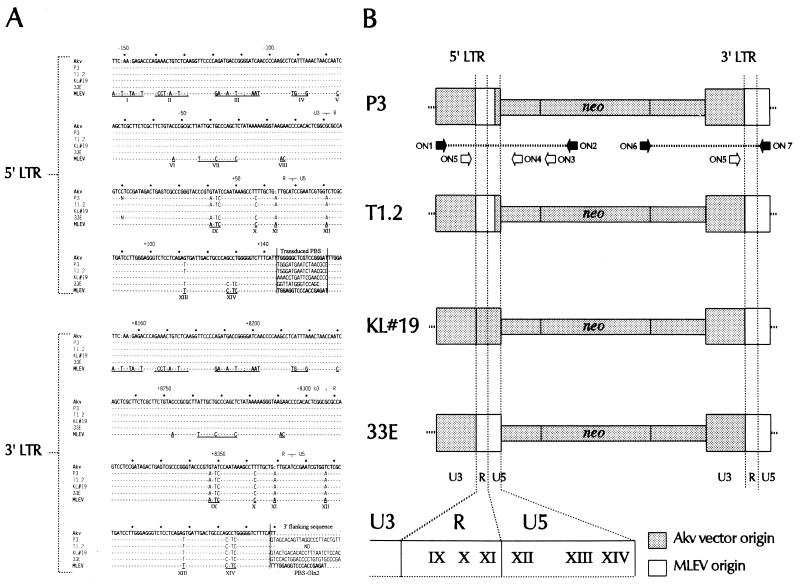
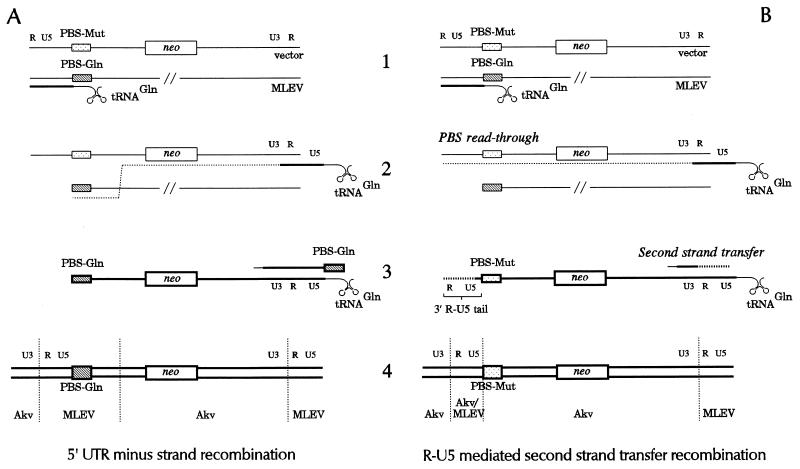
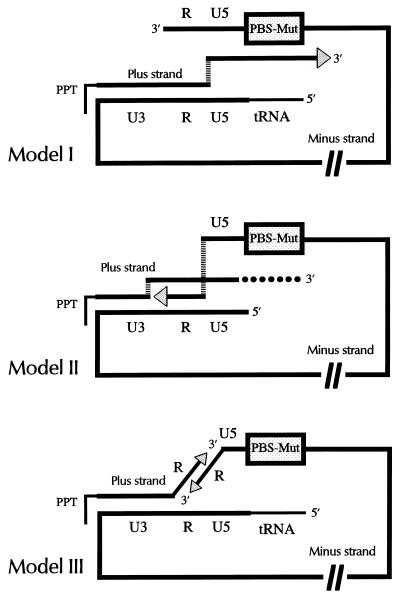
We have previously demonstrated recombinational rescue of primer binding site (PBS)-impaired Akv murine leukemia virus-based vectors involving initial priming on endogenous viral sequences and template switching during cDNA synthesis to obtain PBS complementarity in second-strand transfer of reverse transcription (Mikkelsen et al., J. Virol. 70:1439-1447, 1996). By use of the same forced recombination system, we have now found recombinant proviruses of different structures, suggesting that PBS knockout vectors may be rescued through initial priming on endogenous virus RNA, read-through of the mutated PBS during minus-strand synthesis, and subsequent second-strand transfer mediated by the R-U5 complementarity of the plus strand and the extended minus-strand DNA acceptor template. Mechanisms for R-U5-mediated second-strand transfer and its possible role in retrovirus replication and evolution are discussed.
Figures
Similar articles
-
Recombination in the 5' leader of murine leukemia virus is accurate and influenced by sequence identity with a strong bias toward the kissing-loop dimerization region.J Virol. 1998 Sep;72(9):6967-78. doi: 10.1128/JVI.72.9.6967-6978.1998. J Virol. 1998. PMID: 9696788 Free PMC article.
-
A preferred region for recombinational patch repair in the 5' untranslated region of primer binding site-impaired murine leukemia virus vectors.J Virol. 1996 Mar;70(3):1439-47. doi: 10.1128/JVI.70.3.1439-1447.1996. J Virol. 1996. PMID: 8627661 Free PMC article.
-
Utilization of nonhomologous minus-strand DNA transfer to generate recombinant retroviruses.J Virol. 1997 Mar;71(3):2487-94. doi: 10.1128/JVI.71.3.2487-2494.1997. J Virol. 1997. PMID: 9032388 Free PMC article.
-
Strand transfer events during HIV-1 reverse transcription.Virus Res. 2008 Jun;134(1-2):19-38. doi: 10.1016/j.virusres.2007.12.017. Epub 2008 Feb 14. Virus Res. 2008. PMID: 18279992 Review.
-
Interaction of retroviral reverse transcriptase with template-primer duplexes during replication.Prog Nucleic Acid Res Mol Biol. 1998;58:339-93. doi: 10.1016/s0079-6603(08)60041-0. Prog Nucleic Acid Res Mol Biol. 1998. PMID: 9308371 Review.
Cited by
-
Recombination in the 5' leader of murine leukemia virus is accurate and influenced by sequence identity with a strong bias toward the kissing-loop dimerization region.J Virol. 1998 Sep;72(9):6967-78. doi: 10.1128/JVI.72.9.6967-6978.1998. J Virol. 1998. PMID: 9696788 Free PMC article.
-
Selection of functional tRNA primers and primer binding site sequences from a retroviral combinatorial library: identification of new functional tRNA primers in murine leukemia virus replication.Nucleic Acids Res. 2000 Feb 1;28(3):791-9. doi: 10.1093/nar/28.3.791. Nucleic Acids Res. 2000. PMID: 10637332 Free PMC article.
-
Duplication of the primary encapsidation and dimer linkage region of human immunodeficiency virus type 1 RNA results in the appearance of monomeric RNA in virions.J Virol. 2001 Mar;75(6):2557-65. doi: 10.1128/JVI.75.6.2557-2565.2001. J Virol. 2001. PMID: 11222678 Free PMC article.
-
Retroviral RNA Dimerization: From Structure to Functions.Front Microbiol. 2018 Mar 22;9:527. doi: 10.3389/fmicb.2018.00527. eCollection 2018. Front Microbiol. 2018. PMID: 29623074 Free PMC article. Review.
-
Effect of distance between homologous sequences and 3' homology on the frequency of retroviral reverse transcriptase template switching.J Virol. 1999 Oct;73(10):7923-32. doi: 10.1128/JVI.73.10.7923-7932.1999. J Virol. 1999. PMID: 10482539 Free PMC article.
References
-
- Ben-Artzi H, Shemesh J, Zeelon E, Amit B, Kleiman L, Gorecki M, Panet A. Molecular analysis of the second template switch during reverse transcription of the HIV RNA template. Biochemistry. 1996;35:10549–10557. - PubMed
-
- Berkhout B, van Wamel J, Klaver B. Requirements for DNA strand transfer during reverse transcription in mutant HIV-1 virions. J Mol Biol. 1995;252:59–69. - PubMed
-
- Champoux J J. Roles of ribonuclease H in reverse transcription. In: Skalka A M, Goff S P, editors. Reverse transcriptase. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory; 1993. pp. 103–117.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
