

doi: 10.1128/JVI.72.3.2548-2553.1998.
The glycoprotein D (US6) homolog is not essential for oncogenicity or horizontal transmission of Marek's disease virus
Affiliations
- PMID: 9499123
- PMCID: PMC109562
- DOI: 10.1128/JVI.72.3.2548-2553.1998
Item in Clipboard
The glycoprotein D (US6) homolog is not essential for oncogenicity or horizontal transmission of Marek's disease virus
J Virol.
1998 Mar.
Abstract
RB1BUS6lacgpt, a Marek's disease virus (MDV) mutant having a disrupted glycoprotein D (gD) homolog gene, established infection and induced tumors in chickens exposed to it by inoculation or by contact. Lymphoblastoid cell lines derived from RB1BUS6lacgpt-induced tumors harbored only the mutant virus. These results provide strong evidence that an intact gD homolog gene is not essential for oncogenicity or horizontal transmission of MDV.
Figures
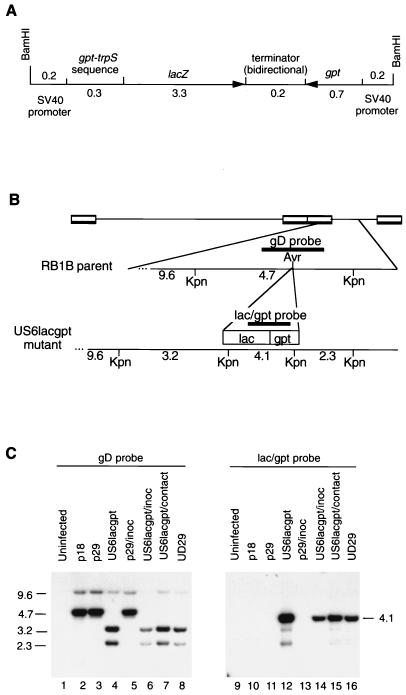
(A) Diagram of the 4.9-kb mutagenesis cassette containing both the lacZ and gpt genes of E. coli. These genes are expressed from simian virus 40 (SV40) early promoter sequences and share a bidirectional transcription terminator. (B) Diagram of the relevant region of the MDV genome for the RB1B parent and the RB1BUS6lacgpt mutant. The positions of the gD-specific and lacZ-specific probes, as well as the KpnI sites, are shown. (C) Southern hybridization analysis of the RB1BUS6lacgpt mutant and corresponding parent strains. Approximately 10 μg of DNA from MDV-infected CEF was KpnI digested, and the resulting fragments were separated on a 0.6% agarose gel and transferred to nitrocellulose. The MDV gD-specific probe used for the Southern analysis was a 1-kb AvaI fragment purified from pMD100 (5) which traverses the AvrII insertion site. The cassette-specific probe was a 2.4-kb EcoRV fragment purified from pMD181 which spans the junction between the gpt and lacZ genes. Probe DNA fragments were gel purified and biotin labelled with the Gene Images random-primed DNA labelling kit (United States Biochemical Corp., Cleveland, Ohio). Lanes: 1 and 9, uninfected CEF DNA; 2 and 10, RB1Bp18-infected CEF DNA; 3 and 11, RB1Bp29-infected CEF DNA; 4 and 12, RB1BUS6lacgpt-infected CEF DNA; 5 and 13, DNA from RB1Bp29 reisolated from inoculated chickens onto CEF; 6 and 14, DNA from RB1BUS6lacgpt reisolated from inoculated chickens onto CEF; 7 and 15, DNA from RB1BUS6lacgpt reisolated from contact-exposed chickens onto CEF; 8 and 16, DNA from virus reactived from the MDCC-UD29 lymphoblastoid cell line, a cell line derived from an RB1BUS6lacgpt-induced tumor present in a contact-exposed chicken. Lanes 1 to 8 were probed with the gD-specific probe, and lanes 9 to 16 were probed with the lacZ-gpt cassette-specific probe. Sizes are indicated in kilobase pairs.
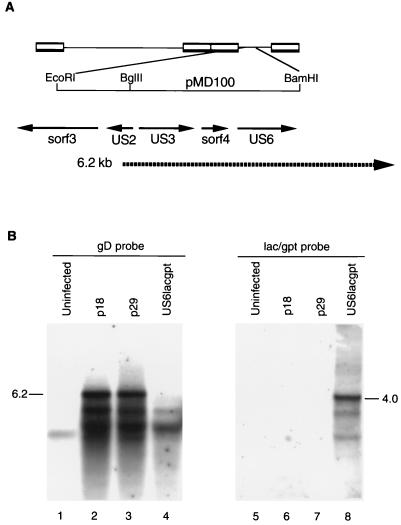
Northern hybridization analysis of RB1BUS6lacgpt mutant and corresponding parent RNAs. Approximately 10 μg of total RNA from each sample was separated on a 1.2% agarose-formaldehyde gel and transferred to nitrocellulose. The gD-specific probe was a T7-derived riboprobe made from pMD168 by in vitro transcription with [32P]CTP. Plasmid pMD168 contains the 0.7-kb AvrII-BamHI fragment from pMD100 (5) and covers the 3′ half of the US6 coding region. The lacZ-specific probe was identical to that used for Southern hybridization. (A) Diagram of the relevant region of the MDV genome showing the positions of known open reading frames and the previously described 6.2-kb transcript that spans the gD coding region. (B) Northern blot analysis. Lanes: 1 and 5, uninfected CEF RNA; 2 and 6, RB1Bp18-infected CEF RNA; 3 and 7, RB1Bp29-infected CEF RNA; 4 and 8, RB1BUS6lacgpt-infected CEF RNA. Lanes 1 to 4 were probed with the gD-specific probe, and lanes 5 to 8 were probed with the lacZ-gpt cassette-specific probe. The positions of the 6.2-kb transcript and the 4.0-kb lacZ-specific transcript are indicated.
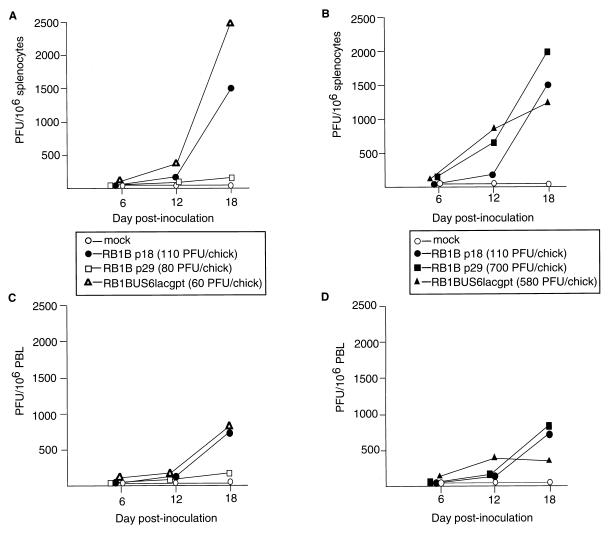
Virus reisolations from chickens inoculated with the RB1BUS6lacgpt and corresponding parent strains. Shown are results of virus reisolations from splenocytes (A and B) or PBL (C and D) of chickens inoculated with mock-infected CEF, RB1Bp18-infected CEF, RB1Bp29-infected CEF, or RB1BUS6lacgpt-infected CEF. Virus doses in PFU per chick are indicated in parentheses. Data from chickens inoculated with RB1Bp29 or RB1BUS6lacgpt at relatively low (A and C) or relatively high doses (B and D) are shown. Data are expressed as PFU obtained on the reisolation dishes per 106 cells.
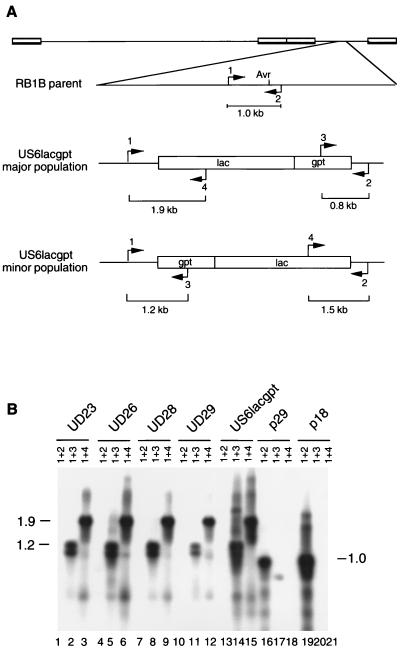
PCR analysis of DNA from lymphoblastoid cell lines derived from RB1BUS6lacgpt-induced tumors. (A) Diagram of the structure of the RB1B parent and both isomers of the RB1BUS6lacgpt mutant with regard to the relevant region of the genome. The orientations of the mutagenesis cassette in the major and minor populations of the mutant are indicated. The positions of PCR primers, as well as the sizes of the predicted products, are shown. Primers 1 and 2, which flank the AvrII insertion site, should direct amplification of a 1.0-kb product from DNA purified from parent-infected CEF but no product from DNA purified from mutant-infected CEF under the PCR conditions used. (B) Southern blot of PCR products from analysis of lymphoblastoid cell line DNAs. PCR products were separated by agarose gel electrophoresis, transferred to nitrocellulose, and probed with the gD-specific probe described in the legend to Fig. 1. The primer pairs used are indicated above the lanes. Templates used are as follows: lanes 1 to 3, MDCC-UD23 DNA; lanes 4 to 6, MDCC-UD26 DNA; lanes 7 to 9, MDCC-UD28 DNA; lanes 10 to 12, MDCC-UD29 DNA; lanes 13 to 15, RB1BUS6lacgpt-infected CEF DNA; lanes 16 to 18, RB1Bp29-infected CEF DNA; lanes 19 to 21, RB1Bp18-infected CEF DNA. Sizes are indicated in kilobase pairs.
Similar articles
-
Characterization of a Marek's disease virus mutant containing a lacZ insertion in the US6 (gD) homologue gene.Virus Genes. 1994 Sep;9(1):5-13. doi: 10.1007/BF01703430. Virus Genes. 1994. PMID: 7871761
-
Horizontal transmission of Marek's disease virus requires US2, the UL13 protein kinase, and gC.J Virol. 2007 Oct;81(19):10575-87. doi: 10.1128/JVI.01065-07. Epub 2007 Jul 18. J Virol. 2007. PMID: 17634222 Free PMC article.
-
The Tegument Protein pUL47 of Marek's Disease Virus Is Necessary for Horizontal Transmission and Is Important for Expression of Glycoprotein gC.J Virol. 2020 Dec 22;95(2):e01645-20. doi: 10.1128/JVI.01645-20. Print 2020 Dec 22. J Virol. 2020. PMID: 32999032 Free PMC article.
-
Latency and tumorigenesis in Marek's disease.Avian Dis. 2013 Jun;57(2 Suppl):360-5. doi: 10.1637/10470-121712-Reg.1. Avian Dis. 2013. PMID: 23901747 Review.
-
T-cell transformation by Marek's disease virus.Trends Microbiol. 1999 Jan;7(1):22-9. doi: 10.1016/s0966-842x(98)01427-9. Trends Microbiol. 1999. PMID: 10068994 Review.
Cited by
-
Transcriptional analysis of Marek's disease virus glycoprotein D, I, and E genes: gD expression is undetectable in cell culture.J Virol. 2001 Mar;75(5):2067-75. doi: 10.1128/JVI.75.5.2067-2075.2001. J Virol. 2001. PMID: 11160711 Free PMC article.
-
Marek's disease virus-1 unique gene LORF1 is involved in viral replication and MDV-1/Md5-induced atrophy of the bursa of Fabricius.PLoS Pathog. 2025 Feb 3;21(2):e1012891. doi: 10.1371/journal.ppat.1012891. eCollection 2025 Feb. PLoS Pathog. 2025. PMID: 39899476 Free PMC article.
-
Clustering of mutations within the inverted repeat regions of a serially passaged attenuated gallid herpesvirus type 2 strain.Virus Genes. 2008 Aug;37(1):69-80. doi: 10.1007/s11262-008-0242-0. Epub 2008 May 31. Virus Genes. 2008. PMID: 18516669
-
Identification of non-essential loci within the Meleagrid herpesvirus 1 genome.Virol J. 2015 Aug 27;12:130. doi: 10.1186/s12985-015-0362-9. Virol J. 2015. PMID: 26307059 Free PMC article.
-
Latest Insights into Unique Open Reading Frames Encoded by Unique Long (UL) and Short (US) Regions of Marek's Disease Virus.Viruses. 2021 May 25;13(6):974. doi: 10.3390/v13060974. Viruses. 2021. PMID: 34070255 Free PMC article. Review.
References
-
- Audonnet J-C, Winslow J, Allen G, Paoletti E. Equine herpesvirus type 1 unique short fragment encodes glycoproteins with homology to herpes simplex virus type 1 gD, gI and gE. J Gen Virol. 1991;71:2969–2978. - PubMed
-
- Buckmaster A E, Scott S D, Sanderson M J, Boursnell M E G, Ross N L J, Binns M M. Gene sequence and mapping data from Marek’s disease virus and herpesvirus of turkeys: implication for herpesvirus classification. J Gen Virol. 1988;69:2033–2042. - PubMed
-
- Calnek B W, Witter R L. Marek’s disease. In: Calnek B W, editor. Diseases of poultry. Ames: Iowa State University Press; 1997. pp. 369–413.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
