

Cdc53 is a scaffold protein for multiple Cdc34/Skp1/F-box proteincomplexes that regulate cell division and methionine biosynthesis in yeast
- PMID: 9499404
- PMCID: PMC316590
- DOI: 10.1101/gad.12.5.692
Cdc53 is a scaffold protein for multiple Cdc34/Skp1/F-box proteincomplexes that regulate cell division and methionine biosynthesis in yeast
Erratum in
- Genes Dev 1998 Oct 1;12(19):3144
Abstract
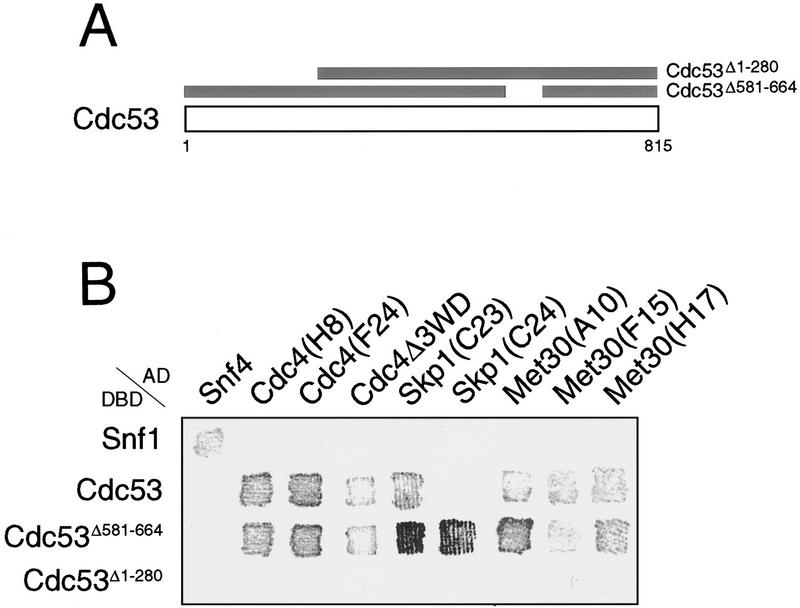
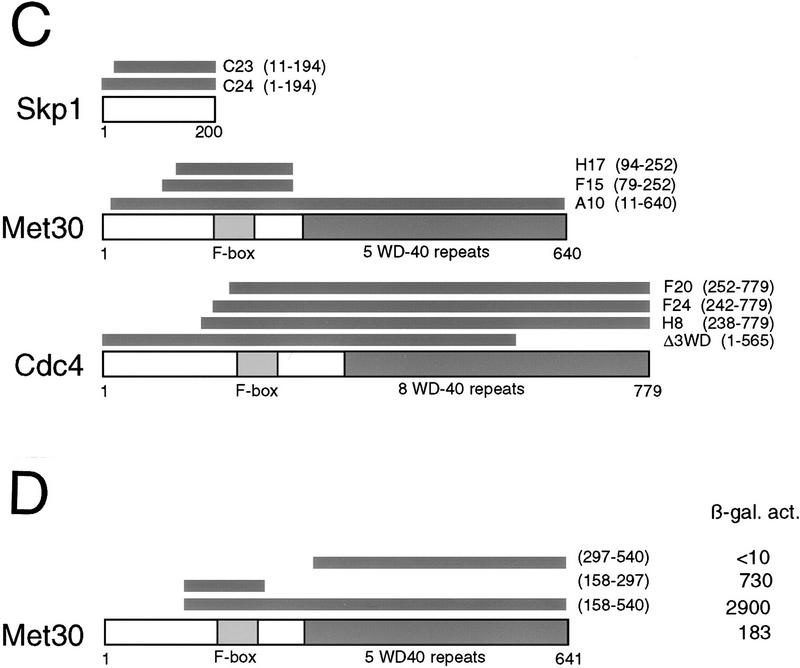
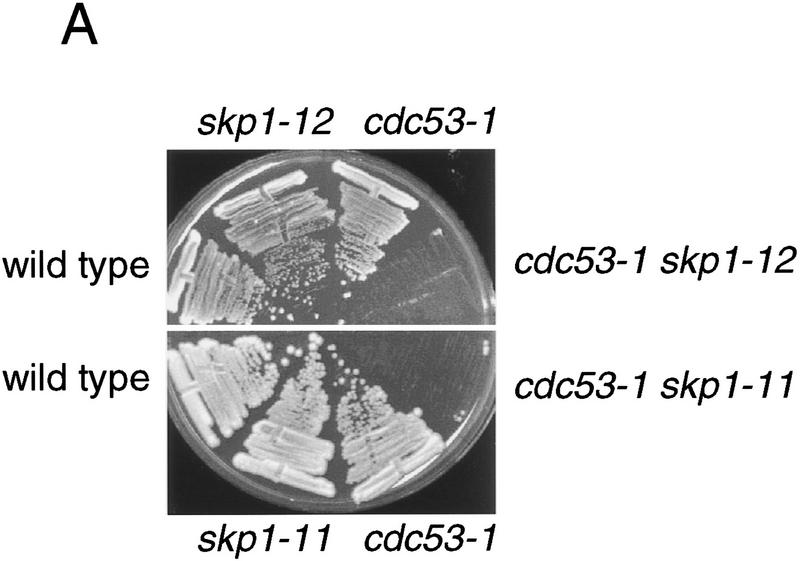
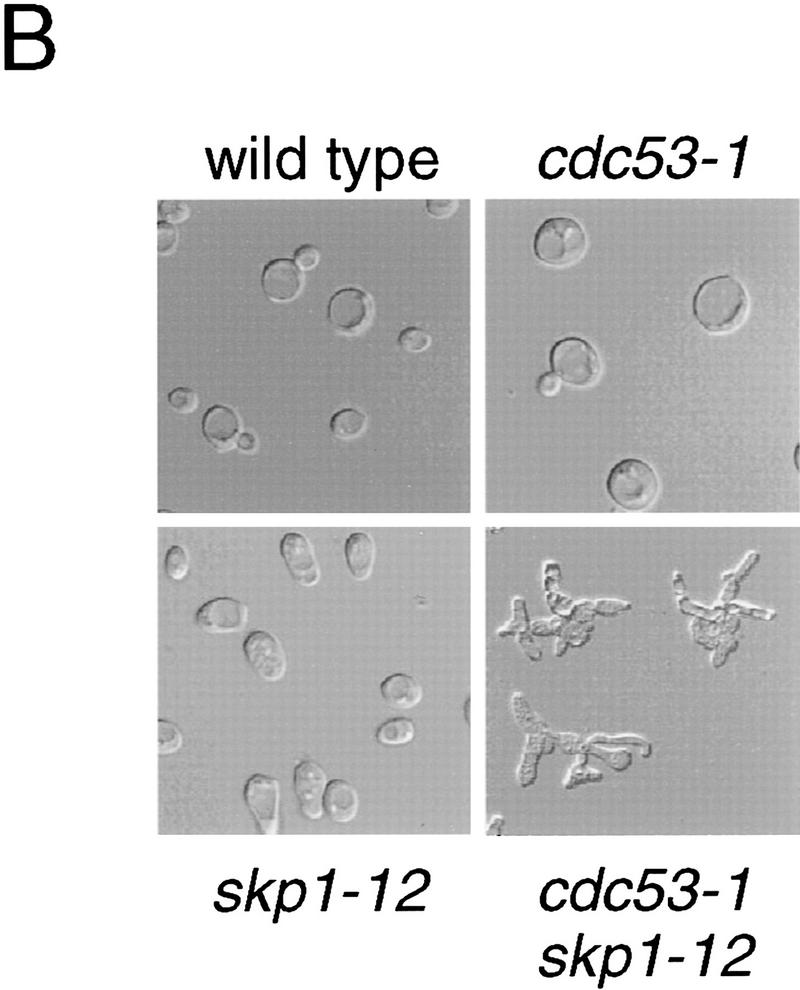
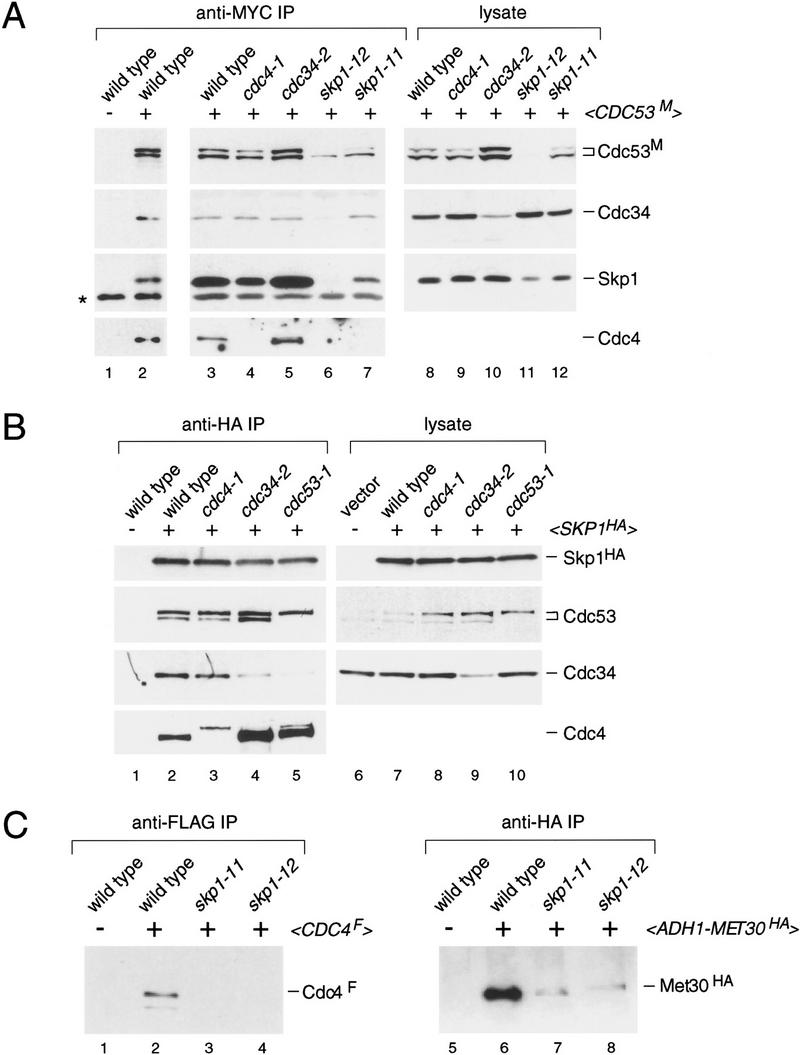
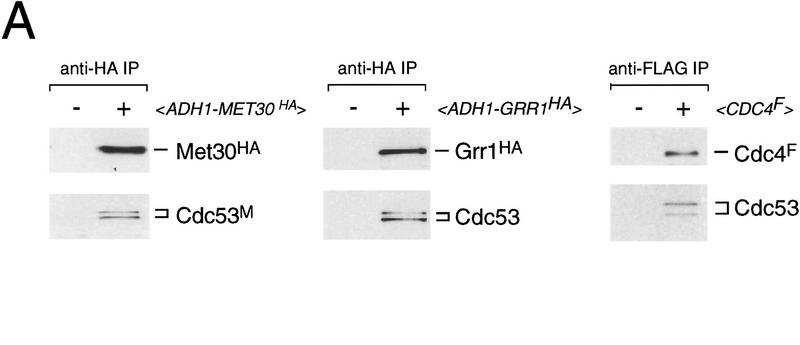
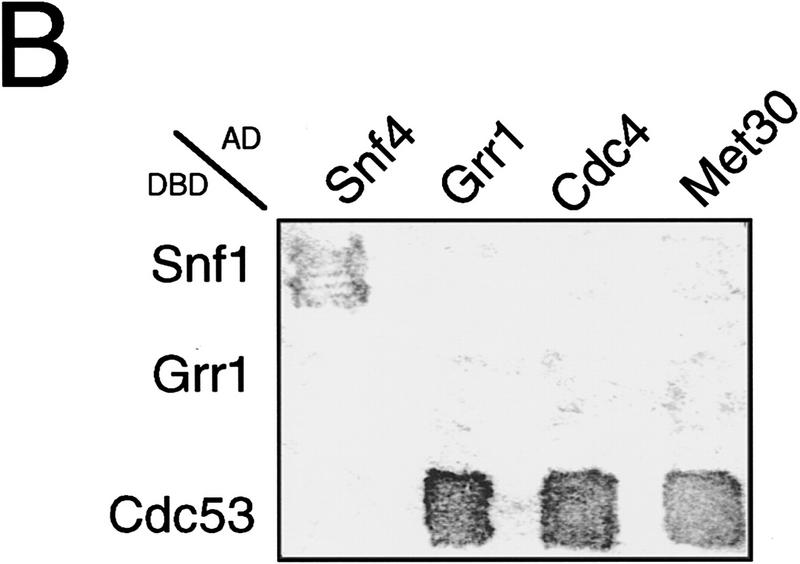
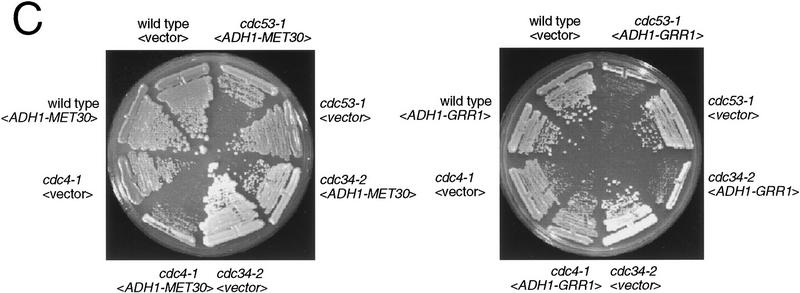
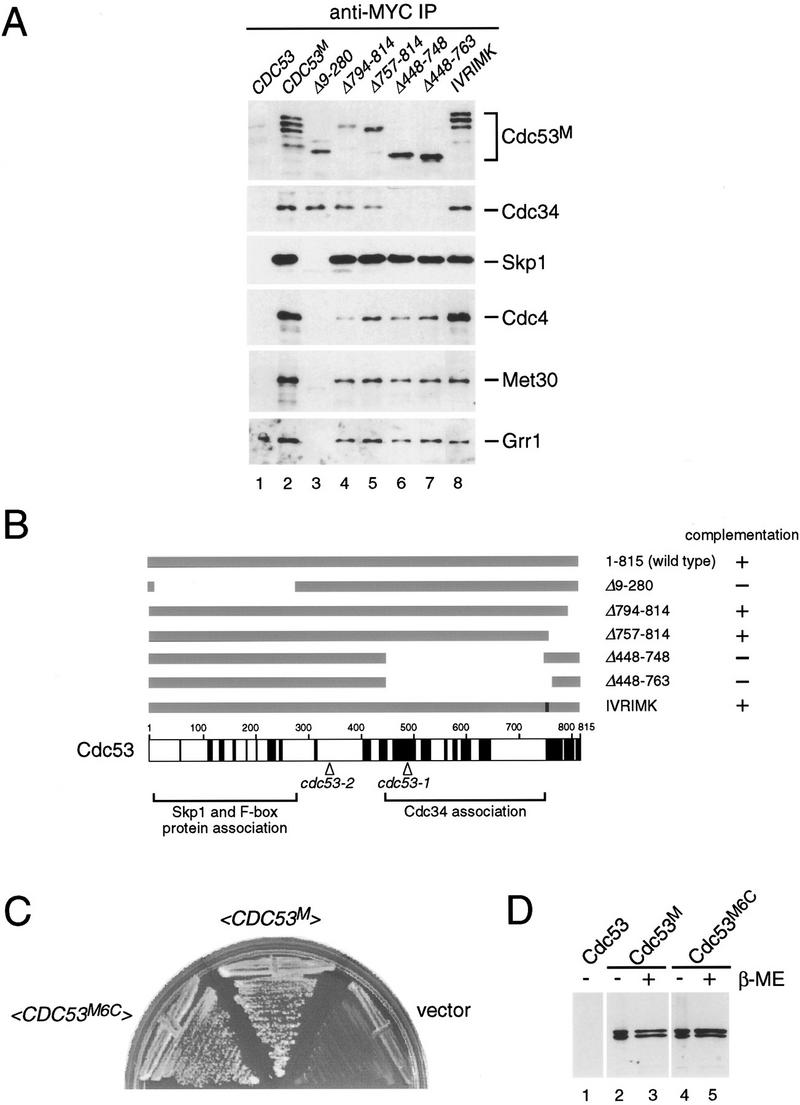
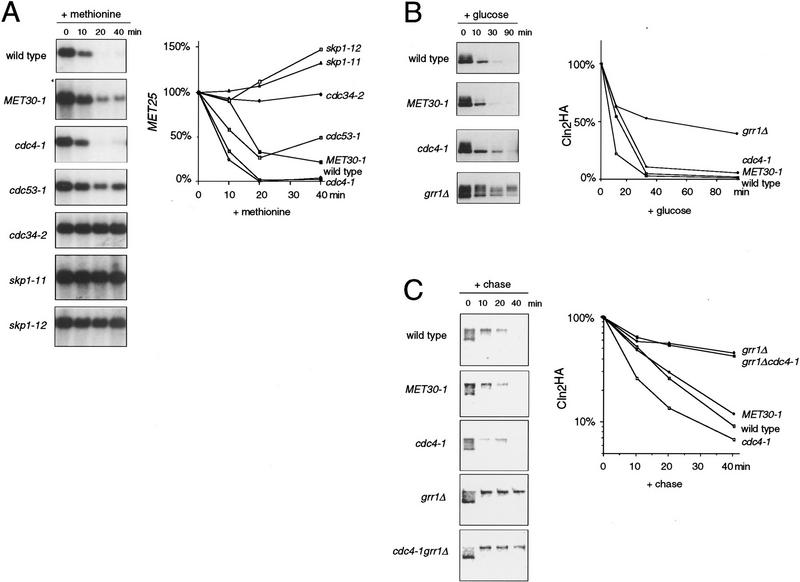
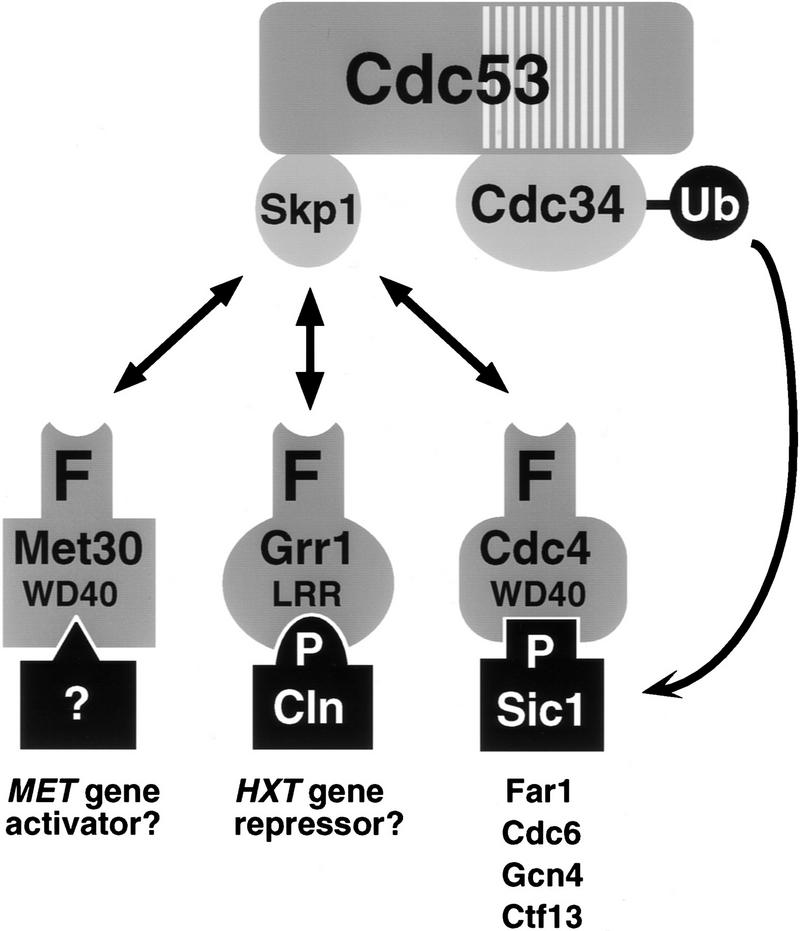
In budding yeast, ubiquitination of the cyclin-dependent kinase (Cdk) inhibitor Sic1 is catalyzed by the E2 ubiquitin conjugating enzyme Cdc34 in conjunction with an E3 ubiquitin ligase complex composed of Skp1, Cdc53 and the F-box protein, Cdc4 (the SCFCdc4 complex). Skp1 binds a motif called the F-box and in turn F-box proteins appear to recruit specific substrates for ubiquitination. We find that Skp1 interacts with Cdc53 in vivo, and that Skp1 bridges Cdc53 to three different F-box proteins, Cdc4, Met30, and Grr1. Cdc53 contains independent binding sites for Cdc34 and Skp1 suggesting it functions as a scaffold protein within an E2/E3 core complex. F-box proteins show remarkable functional specificity in vivo: Cdc4 is specific for degradation of Sic1, Grr1 is specific for degradation of the G1 cyclin Cln2, and Met30 is specific for repression of methionine biosynthesis genes. In contrast, the Cdc34-Cdc53-Skp1 E2/E3 core complex is required for all three functions. Combinatorial control of SCF complexes may provide a basis for the regulation of diverse cellular processes.
Figures
References
-
- Amon A, Irniger S, Nasmyth K. Closing the cell cycle circle in yeast: G2 cyclin proteolysis initiated at mitosis persists until the activation of G1 cyclins in the next cycle. Cell. 1994;77:1037–1050. - PubMed
-
- Bai C, Sen P, Hofmann K, Ma L, Goebl M, Harper JW, Elledge SJ. SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box. Cell. 1996;86:263–274. - PubMed
-
- Barral Y, Jentsch S, Mann C. G1 cyclin turnover and nutrient uptake are controlled by a common pathway in yeast. Genes & Dev. 1995;9:399–409. - PubMed
-
- Blondel M, Mann C. G2 cyclins are required for the degradation of G1 cyclins in yeast. Nature. 1996;384:279–282. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials