

An embryo-defective mutant of arabidopsis disrupted in the final step of biotin synthesis
- PMID: 9501126
- PMCID: PMC35095
- DOI: 10.1104/pp.116.3.935
An embryo-defective mutant of arabidopsis disrupted in the final step of biotin synthesis
Abstract
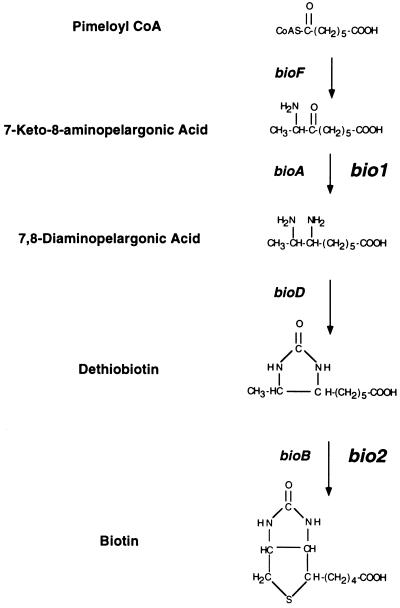
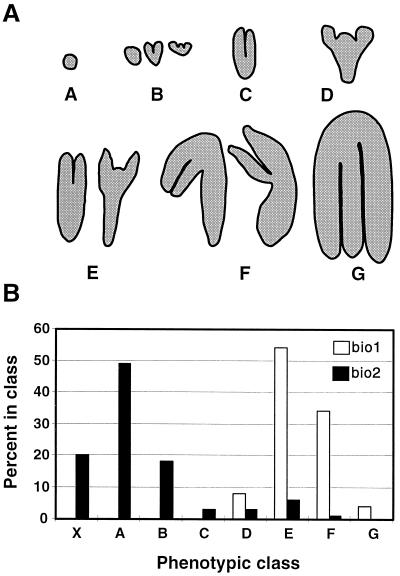
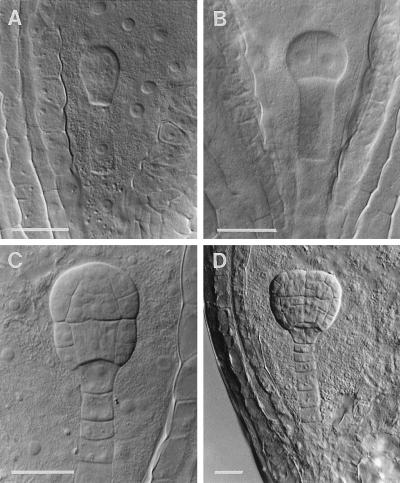
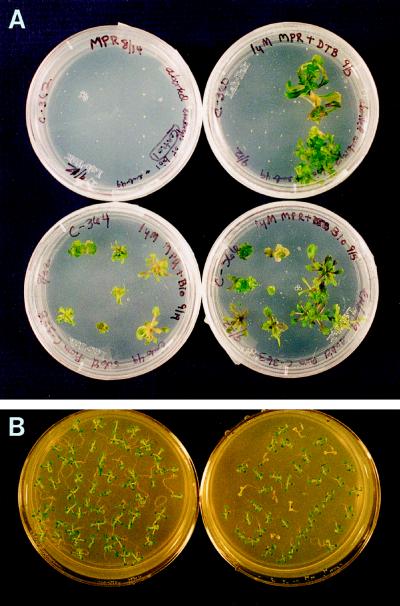
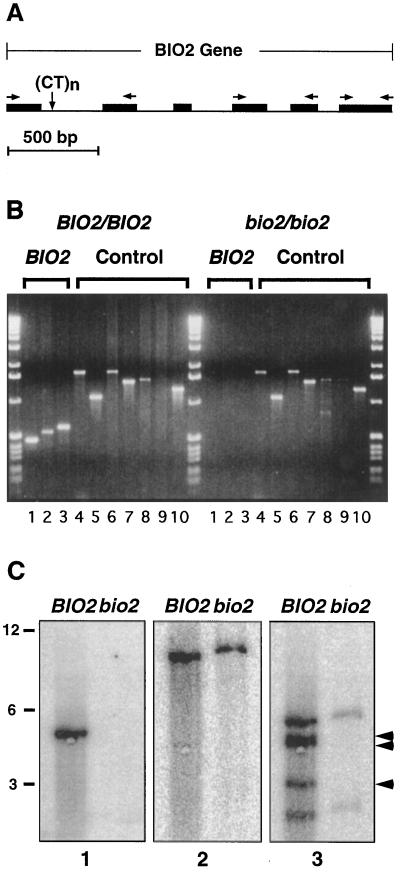
Auxotrophic mutants have played an important role in the genetic dissection of biosynthetic pathways in microorganisms. Equivalent mutants have been more difficult to identify in plants. The bio1 auxotroph of Arabidopsis thaliana was shown previously to be defective in the synthesis of the biotin precursor 7, 8-diaminopelargonic acid. A second biotin auxotroph of A. thaliana has now been identified. Arrested embryos from this bio2 mutant are defective in the final step of biotin synthesis, the conversion of dethiobiotin to biotin. This enzymatic reaction, catalyzed by the bioB product (biotin synthase) in Escherichia coli, has been studied extensively in plants and bacteria because it involves the unusual addition of sulfur to form a thiophene ring. Three lines of evidence indicate that bio2 is defective in biotin synthase production: mutant embryos are rescued by biotin but not dethiobiotin, the mutant allele maps to the same chromosomal location as the cloned biotin synthase gene, and gel-blot hybridizations and polymerase chain reaction amplifications revealed that homozygous mutant plants contain a deletion spanning the entire BIO2-coding region. Here we describe how the isolation and characterization of this null allele have provided valuable insights into biotin synthesis, auxotrophy, and gene redundancy in plants.
Figures
References
-
- Alexeev D, Bury SM, Boys CWG, Turner MA, Sawyer L, Ramsey AJ, Baxter HC, Baxter RL. Sequence and crystallization of Escherichia coli dethiobiotin synthetase, the penultimate enzyme of biotin biosynthesis. J Mol Biol. 1994;235:774–776. - PubMed
-
- Ashburner M. Drosophila: A Laboratory Handbook. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989.
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. New York: John Wiley & Sons; 1987.
-
- Baldet P, Alban C, Axiotis S, Douce R. Localization of free and bound biotin in cells from green pea leaves. Arch Biochem Biophys. 1993a;303:67–73. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
