

Differential expression of a novel gene in response to coronatine, methyl jasmonate, and wounding in the Coi1 mutant of Arabidopsis
- PMID: 9501136
- PMCID: PMC35073
- DOI: 10.1104/pp.116.3.1037
Differential expression of a novel gene in response to coronatine, methyl jasmonate, and wounding in the Coi1 mutant of Arabidopsis
Abstract
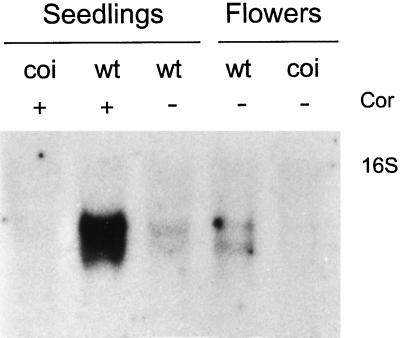
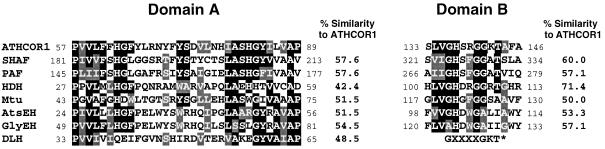
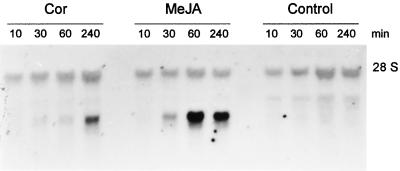
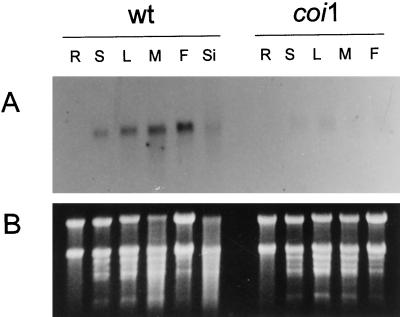
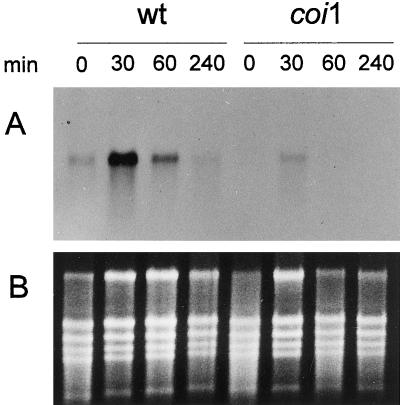
Coronatine is a phytotoxin produced by some plant-pathogenic bacteria. It has been shown that coronatine mimics the action of methyl jasmonate (MeJA) in plants. MeJA is a plant-signaling molecule involved in stress responses such as wounding and pathogen attack. In Arabidopsis thaliana, MeJA is essential for pollen grain development. The coi1 (for coronatine-insensitive) mutant of Arabidopsis, which is insensitive to coronatine and MeJA, produces sterile male flowers and shows an altered response to wounding. When the differential display technique was used, a message that was rapidly induced by coronatine in wild-type plants but not in coi1 was identified and the corresponding cDNA was cloned. The coronatine-induced gene ATHCOR1 (for A. thaliana coronatine-induced) is expressed in seedlings, mature leaves, flowers, and siliques but was not detected in roots. The expression of this gene was dramatically reduced in coi1 plants, indicating that COI1 affects its expression. ATHCOR1 was rapidly induced by MeJA and wounding in wild-type plants. The sequence of ATHCOR1 shows no strong homology to known proteins. However, the predicted polypeptide contains a conserved amino acid sequence present in several bacterial, animal, and plant hydrolases and includes a potential ATP/GTP-binding-site motif (P-loop).
Figures

References
-
- Bender CL, Stone HE, Sims JJ, Cookey DA. Reduced pathogen fitness of Pseudomonas syringae pv tomato Tn5 mutants defective in coronatine production. Physiol Mol Plant Pathol. 1987;30:273–283.
-
- Beveridge AJ, Ollis DL. A theoretical study of substrate-induced activation of dienelactone hydrolase. Protein Eng. 1995;8:135–142. - PubMed
-
- Blée E, Schuber F. Biosynthesis of cutin monomers: involvement of a lipoxygenase/peroxygenase pathway. Plant J. 1993;4:113–123.
-
- Coehn Y, Gisi U, Niderman T. Local and systemic protection against Phytophthora infestans induced in potato and tomato plants by jasmonic acid and jasmonic methyl ester. Phytopathology. 1993;83:1054–1062.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
