

The role of iron-deficiency stress responses in stimulating heavy-metal transport in plants
- PMID: 9501139
- PMCID: PMC35076
- DOI: 10.1104/pp.116.3.1063
The role of iron-deficiency stress responses in stimulating heavy-metal transport in plants
Abstract
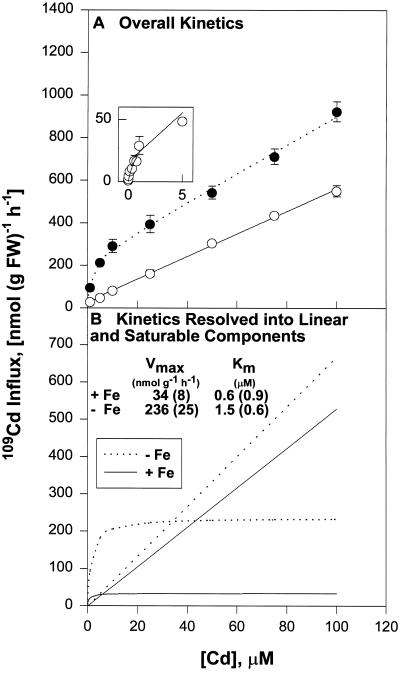
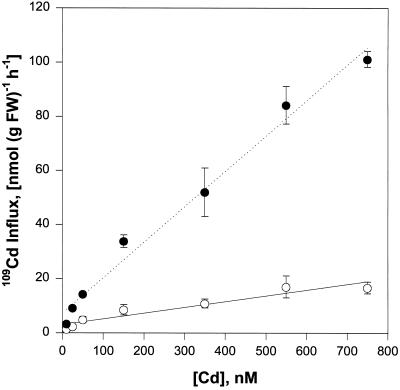
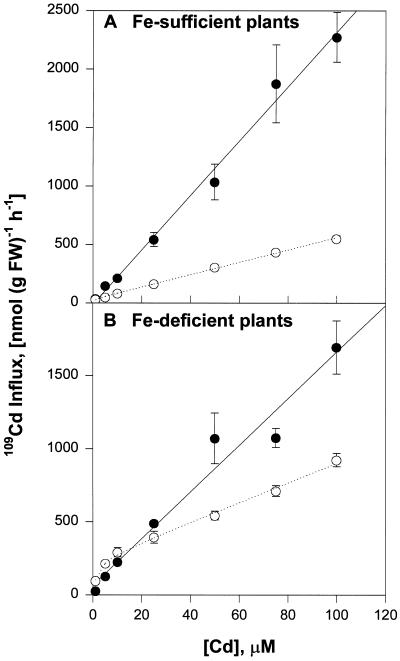
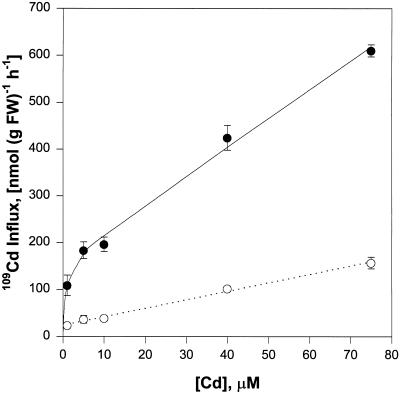
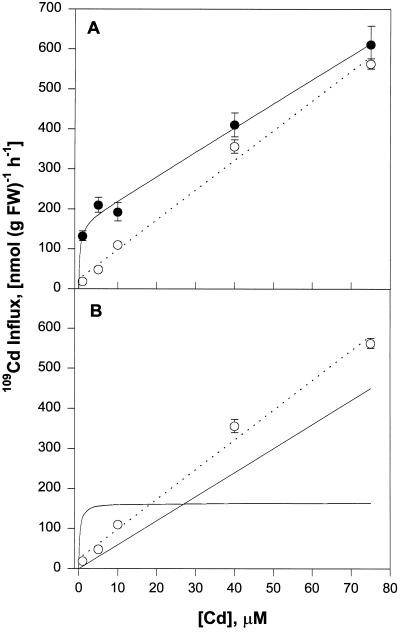
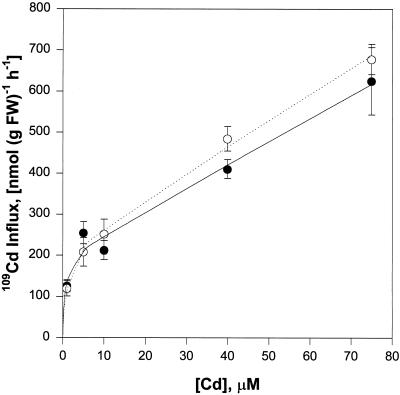
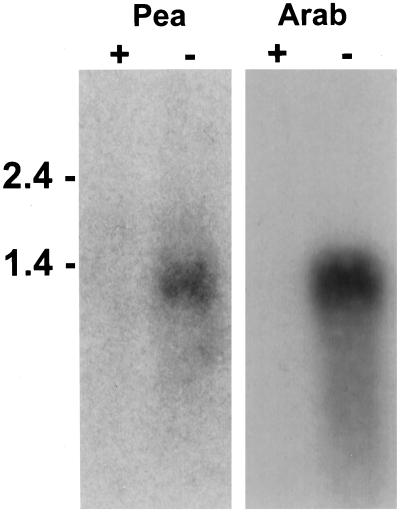
Plant accumulation of Fe and other metals can be enhanced under Fe deficiency. We investigated the influence of Fe status on heavy-metal and divalent-cation uptake in roots of pea (Pisum sativum L. cv Sparkle) seedlings using Cd2+ uptake as a model system. Radiotracer techniques were used to quantify unidirectional 109Cd influx into roots of Fe-deficient and Fe-sufficient pea seedlings. The concentration-dependent kinetics for 109Cd influx were graphically complex and nonsaturating but could be resolved into a linear component and a saturable component exhibiting Michaelis-Menten kinetics. We demonstrated that the linear component was apoplastically bound Cd2+ remaining in the root cell wall after desorption, whereas the saturable component was transporter-mediated Cd2+ influx across the root-cell plasma membrane. The Cd2+ transport system in roots of both Fe-deficient and Fe-sufficient seedlings exhibited similar Michaelis constant values, 1.5 and 0.6 m, respectively, for saturable Cd2+ influx, whereas the maximum initial velocity for Cd2+ uptake in Fe-deficient seedlings was nearly 7-fold higher than that in Fe-grown seedlings. Investigations into the mechanistic basis for this response demonstrated that Fe-deficiency-induced stimulation of the plasma membrane H+-ATPase did not play a role in the enhanced Cd2+ uptake. Expression studies with the Fe2+ transporter cloned from Arabidopsis, IRT1, indicated that Fe deficiency induced the expression of this transporter, which might facilitate the transport of heavy-metal divalent cations such as Cd2+ and Zn2+, in addition to Fe2+.
Figures
References
-
- Ambler JE, Brown JC, Gauch HG. Sites of iron reduction in soybean plants. Agron J. 1971;63:95–97.
-
- Askwith C, Eide D, Van Ho A, Bernard PS, Li L, Davis-Kaplan S, Sipe DM, Kaplan J. The FET3 gene of S. cerevisiae encodes a multicopper oxidase required for ferrous iron uptake. Cell. 1994;76:403–410. - PubMed
-
- Bienfait HF. Mechanisms in Fe-efficiency reactions of higher plants. J Plant Nutr. 1988;11:605–629.
-
- Bienfait HF, Bino RJ, van der Bliek AM, Duivenvoorden JF, Fontain JM. Characterization of ferric reducing activity in roots of Fe-deficient Phaseolus vulgaris. Physiol Plant. 1983;59:196–202.
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
