

Fluctuations in cyclin E levels are required for multiple rounds of endocycle S phase in Drosophila
- PMID: 9501987
- PMCID: PMC2754236
- DOI: 10.1016/s0960-9822(98)70089-2
Fluctuations in cyclin E levels are required for multiple rounds of endocycle S phase in Drosophila
Abstract
The precise cell-cycle alternation of S phase and mitosis is controlled by alternating competence of nuclei to respond to S-phase-inducing factors [1]. Nuclei acquire competence to replicate at the low point in cyclin-dependent kinase (Cdk) activities that follows mitotic destruction of cyclins. The elevation of Cdk activity late in G1 is thought to drive cells into S phase and to block replicated DNA from re-acquiring replication competence [2]. Whereas mitosis is normally required to eliminate the cyclins prior to another cycle of replication, experimental elimination of Cdk activity in G2 can restore competence to replicate [3-6]. Here, we examine the roles of Cdks in the endocycies of Drosophila [7]. In these cycles, rounds of discrete S phases without intervening mitoses result in polyteny. Cyclins A and B are lost in cells as they enter endocycles [8,9], and pulses of Cyclin E expression drive endocycle S phases [10-12]. To address whether oscillations of Cyclin E expression are required for endocycles, we expressed Cyclin E continuously in Drosophila salivary glands. Growth of the cells was severely inhibited, and a period of DNA replication was induced but further replication was inhibited. This replication inhibition could be overcome by the kinase inhibitor 6-dimethylaminopurine (6-DMAP), but not by expression of subunits of the transcription factor E2F. These results indicate that endocycle S phases require oscillations in Cdk activity, but, in contrast to oscillations in mitotic cells, these occur independently of mitosis.
Figures
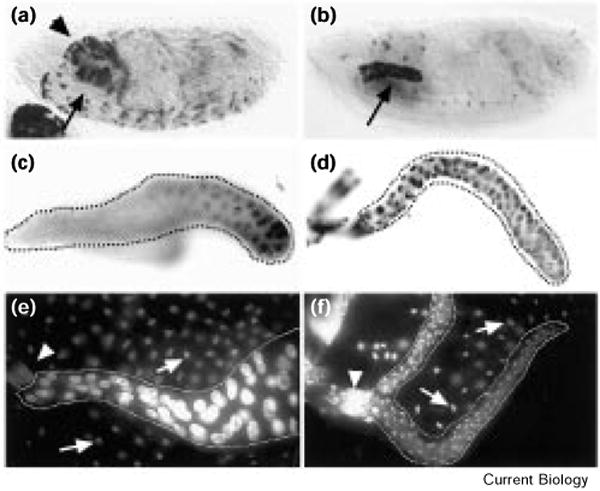
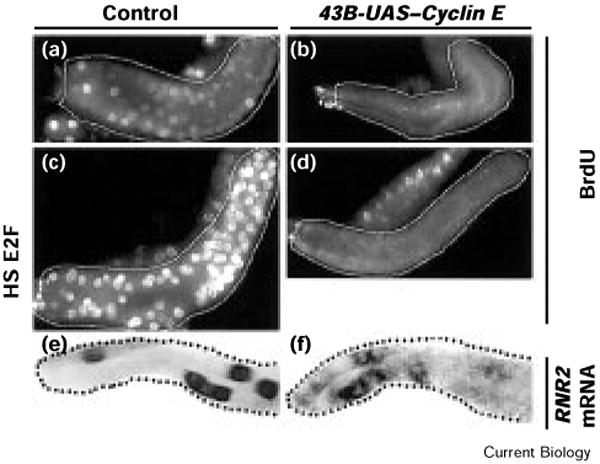
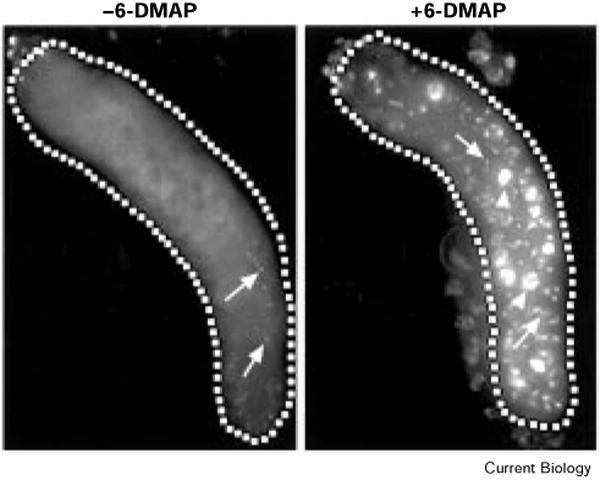
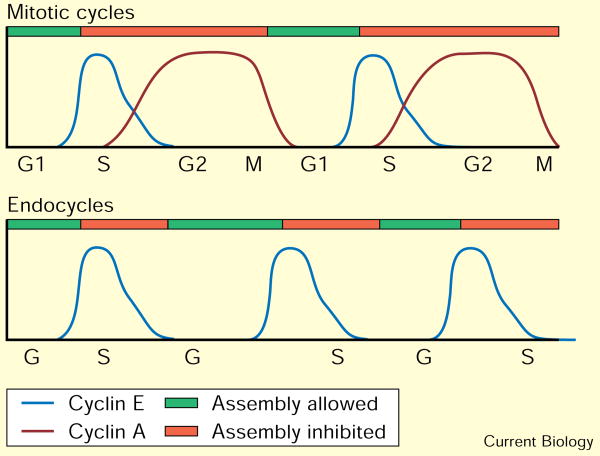
References
-
- Rao PN, Johnson RT. Mammalian cell fusion: studies on the regulation of DNA synthesis and mitosis. Nature. 1970;225:159–164. - PubMed
-
- Broek D, Bartlett R, Crawford K, Nurse P. Involvement of p34cdc2 in establishing the dependency of S phase on mitosis. Nature. 1991;349:388–393. - PubMed
-
- Hayles J, Fisher D, Woollard A, Nurse P. Temporal order of S phase and mitosis in fission yeast is determined by the state of the p34cdc2-mitotic B cyclin complex. Cell. 1994;78:813–822. - PubMed
-
- Dahmann C, Diffley JF, Nasmyth KA. S-phase-promoting cyclin-dependent kinases prevent re-replication by inhibiting the transition of replication origins to a pre-replicative state. Curr Biol. 1995;5:1257–1269. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
