

doi: 10.1523/JNEUROSCI.18-07-02520.1998.
Astrocytic gap junctions remain open during ischemic conditions
Affiliations
- PMID: 9502812
- PMCID: PMC6793088
- DOI: 10.1523/JNEUROSCI.18-07-02520.1998
Item in Clipboard
Astrocytic gap junctions remain open during ischemic conditions
J Neurosci.
.
Abstract
Gap junctions are highly conductive channels that allow the direct transfer of intracellular messengers such as Ca2+ and inositol triphosphate (IP3) between interconnected cells. In brain, astrocytes are coupled extensively by gap junctions. We found here that gap junctions among astrocytes in acutely prepared brain slices as well as in culture remained open during ischemic conditions. Uncoupling first occurred after the terminal loss of plasma membrane integrity. Gap junctions therefore may link ischemic astrocytes in an evolving infarct with the surroundings. The free exchange of intracellular messengers between dying and potentially viable astrocytes might contribute to secondary expansion of ischemic lesions.
Figures
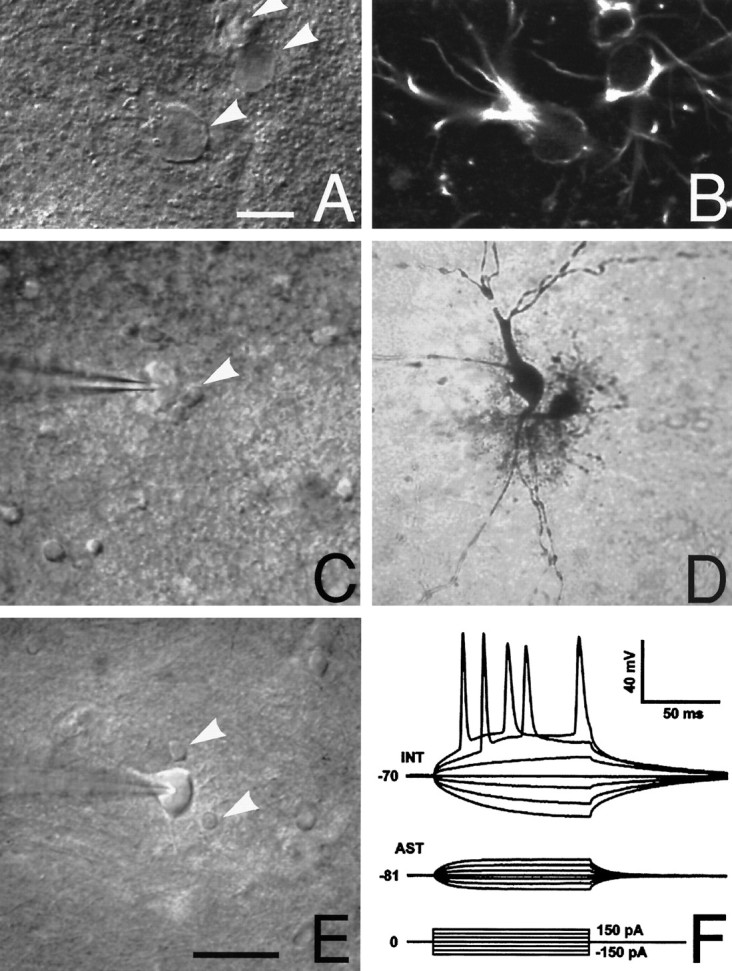
Astrocytes in the stratum radiatum of the hippocampal CA1 region. A, Differential interference contrast (DIC) microscopy was used to identify both astrocytes and neurons before electrophysiological recordings. Astrocytes were identified by their characteristically small rounded cell bodies and poorly defined irregular processes (arrowheads). Scale bar, 10 μm. B, Immunoreactivity against glial fibrillary acidic protein (GFAP) in the same field as A. Three cells identified as astrocytes by DIC microscopy were GFAP-positive. C, Another hippocampal slice visualized by DIC microscopy. Two cells, one with a morphology typical of astrocytes (arrowhead) and one interneuron (impaled by patch electrode), are indicated. D, Biocytin was injected into both of these cells to compare DIC with cellular morphology. The cell identified by DIC as an astrocyte was characterized by an extensive array of branched short processes, phenotypical for astrocytes. In contrast, the presumed interneuron had few, but evenly sized, unbranched processes—a morphology stereotypical for neurons. All eight pairs of biocytin-injected astrocytes and neurons had similar patterns of cellular morphology. E, A representative field containing one interneuron (impaled by patch electrode) surrounded by two representative astrocytes (arrowheads). Scale bar, 50 μm for C–E. F, Astrocytes and neurons also were identified by their electrophysiological properties. Shown are representative tracings from an astrocyte (AST) with a resting membrane potential of −81 mV. No action potentials were evoked by depolarization pulses. All 60 astrocytes identified by DIC microscopy lacked depolarization-evoked action potentials. In contrast, trains of action potentials were evoked by depolarization pulses in an interneuron (INT) with a resting membrane potential of −70 mV.
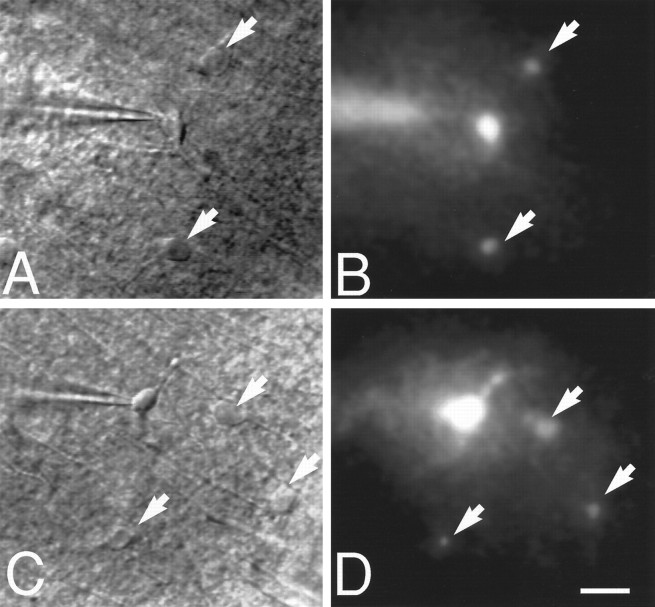
Intercellular injection of Lucifer yellow disclosed that astrocytic gap junctions remain open during ischemic conditions in brain slices. A, A representative field containing three cells with a morphology typical for astrocytes under DIC microscopy. One cell was patched by an electrode; the two remaining cells are indicated by arrows. B, Dye coupling during normoxic control conditions in the same field. Lucifer yellow (4%) was injected into the patched cell (resting membrane potential, −81 mV). After 10 min, several neighboring cells (arrows) were brightly fluorescent, indicating that Lucifer yellow had diffused from the injected cell to surrounding coupled cells. C, A representative field containing four cells with a morphology typical for astrocytes during ischemic conditions. D, Diffusion of Lucifer yellow in the same field. Lucifer yellow was injected into the patched astrocyte 15 min after the induction of ischemia. Ten minutes later, three neighboring cells stained brightly with Lucifer yellow. Thus, astrocytic coupling persisted during ischemic conditions. Intercellular Lucifer yellow diffusion was observed in eight other slices during ischemic conditions. Scale bar, 20 μm.
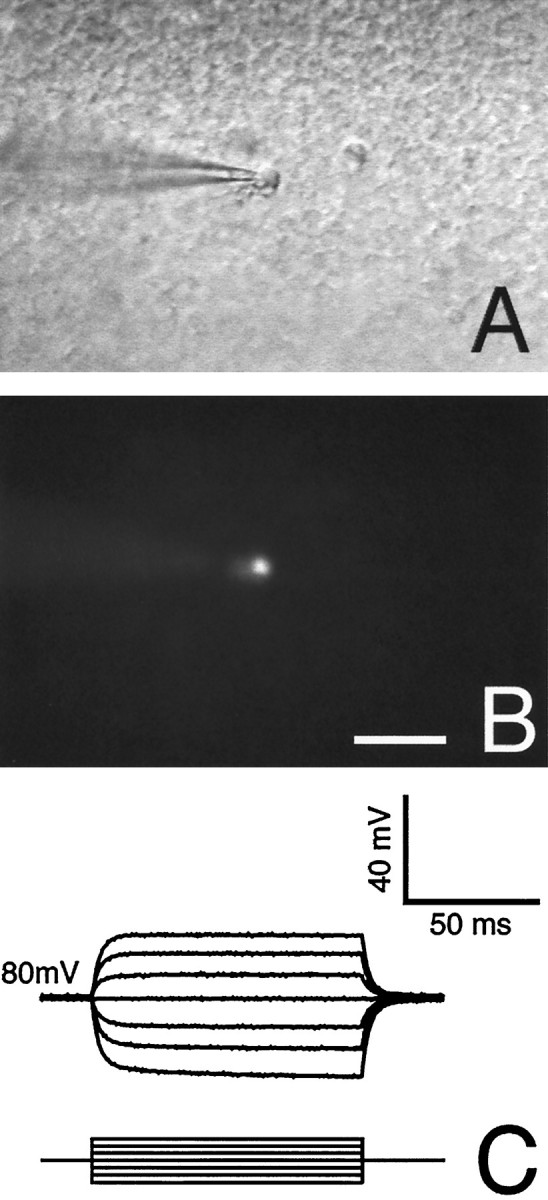
Surface astrocytes in acutely prepared slices do not transfer Lucifer yellow. A, DIC micrograph of a representative field containing two surface cells with a morphology typical for astrocytes (one cell is patched). B, Lucifer yellow was injected into one cell and allowed to diffuse for 10 min during normoxic control conditions. No dye diffusion to surrounding cells was detected. Absence of dye diffusion was observed in all eight injections in surface astrocytes. C, Tracing from the same cell with a resting membrane potential of −80 mV. Input resistance was several orders of magnitude higher than deeper astrocytes (compare with Fig. 1F). Scale bar, 30 μm.
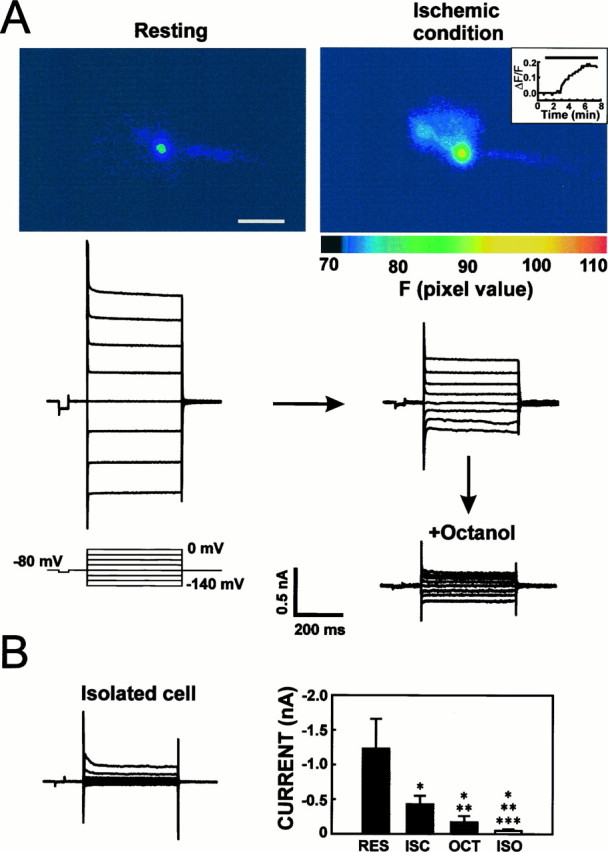
Quantification of gap junction coupling during ischemic conditions in acutely prepared slices. In vitroischemic conditions reduced, but did not block, astrocytic gap junctions in brain slices. A, An astrocyte in stratum radiatum of the hippocampal CA1 region was patched in the whole-cell voltage-clamp configuration, and current was activated from a holding potential of −80 mV (step protocol, see inset). The patch pipette contained the calcium indicator fluo-3. Anoxic aglycemia induced a twofold increase in the emission signal of fluo-3 within 5 min of ischemia. Inset, Map ΔF/F as a function of time in the same astrocyte. Line indicates ischemic conditions. Large currents were activated in the same cell during resting conditions, as expected in a gap junction-coupled cell. Ischemic conditions decreased currents when both depolarization and hyperpolarization steps from +80 to −60 mV were delivered. The observed decrease in activated current occurred concomitantly with the increase in fluo-3 signal. Octanol (5 mm ) decreased the current further, indicating that ischemia reduced, but did not block, astrocytic gap junctions. B, Current recordings from an isolated surface astrocyte revealed an even lower amplitude of activated current than in octanol-treated coupled cells, suggesting that 5 mm octanol did not block gap junction coupling completely. Right panel summarizes the mean current activated when 60 mV hyperpolarization steps were delivered during resting condition (RES), ischemic condition (ISC; 20–40 min), subsequent octanol treatment of the ischemic astrocytes (OCT), and in isolated astrocytes patched at the surface of the slice (ISO). Recordings during control and ischemic conditions as well as during octanol exposure were obtained in the same cells (n = 7), whereas isolated cells were recorded separately (n = 9). Coupling during resting condition and responses to anoxic aglycemia were qualitatively similar in cortical and hippocampal astrocytes, and the data have been pooled. Values are mean ± SD. *Denotes different from control condition; **denotes different from ischemic condition; ***denotes different from octanol treatment at p < 0.001, using Student’st test. Scale bar, 20 μm.
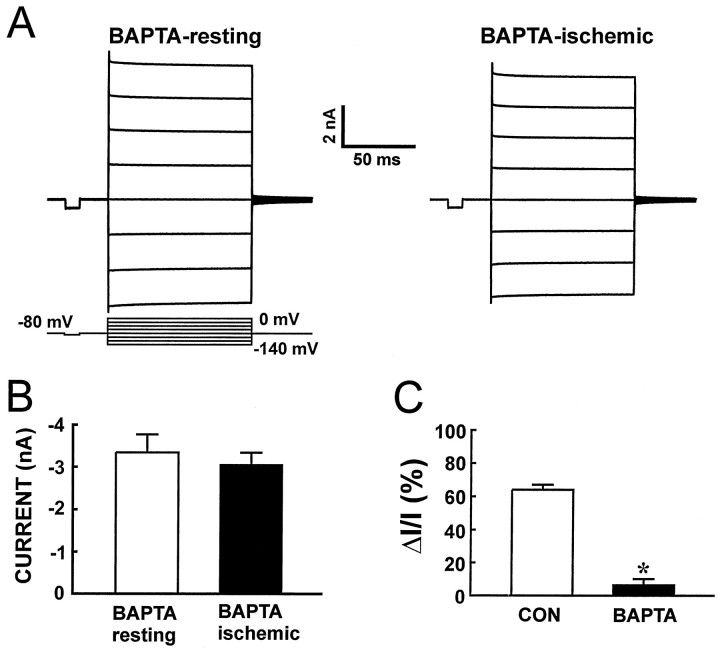
Buffering intracellular Ca2+attenuates the uncoupling effect of ischemia. A, A single astrocyte was patched with an electrode solution containing a 20 mm concentration of the calcium chelator BAPTA. The amplitude of activity-induced current was considerably higher in the presence than in the absence of BAPTA, suggesting that the lowering of [Ca2+]i increased coupling during the resting condition. Anoxic aglycemia was induced 15 min later. In contrast to the previous figure (see Fig. 4), activated current did not decrease in BAPTA-loaded cell recordings during ischemic conditions.B, Summarized amplitude of current activated by a 60 mV hyperpolarization step in BAPTA-loaded astrocytes during resting and ischemic conditions (n = 6). The amplitude of current did not decrease significantly in the presence of BAPTA (paired Student’s t test). C, Comparison of the relative decrease in current (ΔI/I) evoked by ischemia in the absence (CON) and presence of BAPTA in the recording electrode. BAPTA loading significantly reduced the uncoupling effect of ischemia (p < 0.01, Student’st test).
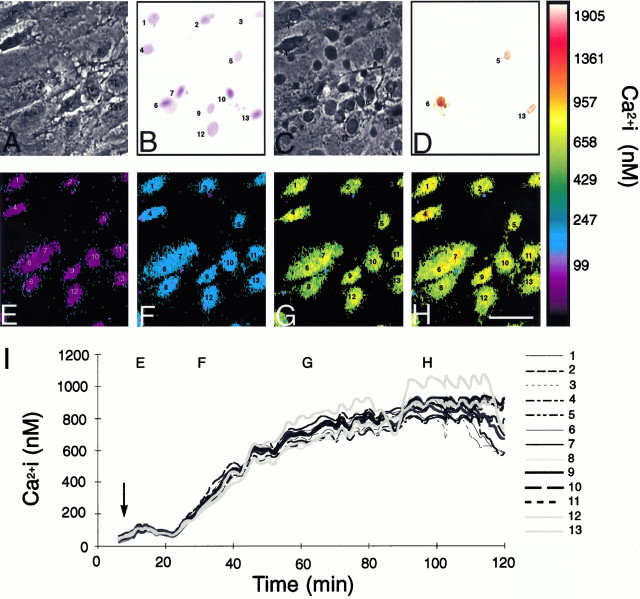
Pattern of astrocytic death after metabolic inhibition during culture conditions. A, Phase-contrast photomicrograph of a representative field of astrocytes during the resting condition. B, The same field after metabolic inhibition. Most cells stained with trypan blue 24 hr after exposure, indicating irreversible loss of viability. C, Corresponding phase-contrast photomicrograph 24 hr after exposure. Cell shrinkage and retraction of processes are evident. D, TUNEL stain of the same field. Only three cells were dUTP-positive.E–H, Pseudocolor display of [Ca2+]i changes in the same field.I, [Ca2+]i changes in individual cells mapped as a function of time. [Ca2+]i increased uniformly in all cells studied. Gap junction function was evaluated in cultures treated similarly (see Fig. 7). The uniform cellular reaction to metabolic inhibition, both with regard to [Ca2+]i changes and irreversible membrane damage, allowed gap junction permeability to be evaluated as a function of both [Ca2+]i and viability. Scale bar, 15 μm.
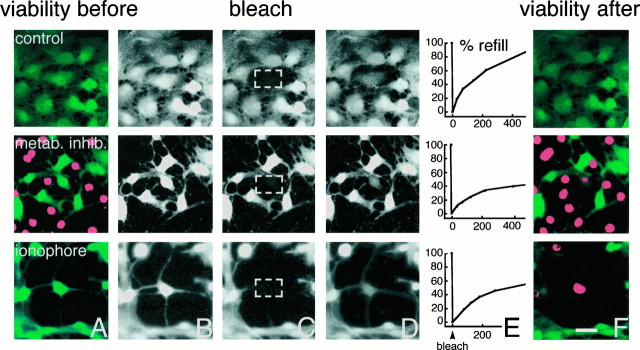
Astrocytic gap junctions remain open during the process of cell death. Cultured astrocytes were loaded with CDCF (green). CDCF is plasma membrane-impermeable, and cell death defined by loss of membrane integrity thus can be detected as a rapid loss of CDCF fluorescence. Concomitantly, propidium iodide (red) gains access to the cell interior and stains the nuclei of dead cells red. Additionally, the small molecular weight of CDCF allows it to diffuse freely across gap junctions so that gap junction permeability can be evaluated by fluorescence recovery after photobleach (FRAP) just before the loss of membrane integrity.Top panels, Fluorescence recovery after photobleach in a healthy culture. A, Fluorescence before photobleach. All cells have intact membranes and therefore do not incorporate propidium iodide but are CDCF-positive (green).B, CDCF fluorescence before photobleach (gray color scale). C, CDCF fluorescence immediately after photobleach. The white square indicates the target area of photobleach.D, The cell has partly regained fluorescence because of influx of CDCF from surrounding gap junction-coupled cells 2 min later.E, Percentage of refill as a function of time in the same cell. Forty percent recovery of fluorescence occurred within 2 min. F, All cells remain viable in this control culture when observed 15 min later (propidium iodide−/CDCF+). Middle panels, Photobleach in a dying culture exposed to KCN and iodoacetate for 80 min. A, Several astrocytes have lost viability in this field (propidium iodide+/CDCF−).B–D, Photobleach in a still viable cell.E, Fluorescence recovered by 21% within 2 min.F, Thirteen minutes later the target cell has lost viability (propidium iodide+/CDCF−). Thus, dying astrocytes remain coupled during the process of cell death. Bottom panels, Photobleach in a dying culture exposed to the calcium ionophore lasalocid 40 min earlier. A, All cells are still propidium iodide−/CDCF+, indicating that no loss of viability has occurred yet. B–D, Photobleach in a still viable cell.E, Fluorescence recovered by 31% within 2 min.F, Seventeen minutes after photobleach, the cell has lost viability (propidium iodide+/CDCF−). Thus, refill occurred minutes before irreversible membrane injury. Scale bar, 15 μm.
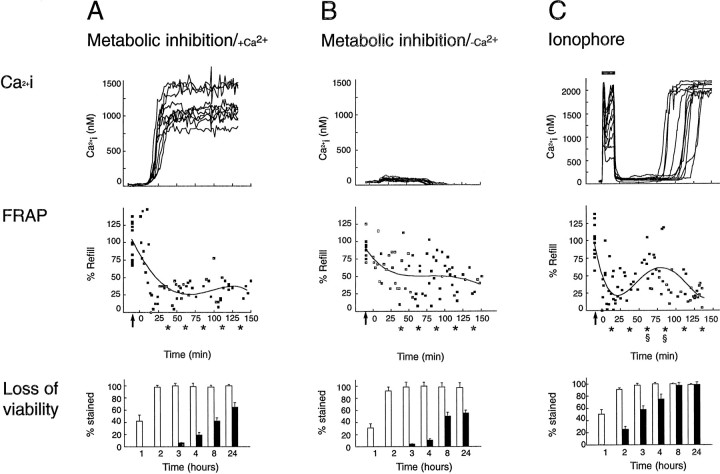
Calcium decreases, but does not block, astrocytic coupling during the process of cell death. Comparison of [Ca2+]i (top panels), gap junction coupling normalized against control values (middle panels), and loss of viability (bottom panels) during and after lethal injury. Control cultures were characterized by a resting [Ca2+]i averaging 50–100 nm and by substantial refill. A, Top panel, A rapid increase in [Ca2+]i and a corresponding decrease in gap junction coupling with no sign of recovery (middle panel) occurred when energy metabolism was inhibited by KCN (inhibitor of oxidation chain) and iodoacetate (blocker of glycolysis). Refill after photobleach in separate experiments is mapped individually (squares) and the data fit by four-order regression. Arrows indicate refill during resting control conditions in the same cultures. Bottom panel, The percentage of cell death after the same procedure. Almost complete loss of viability was observed at 2–3 hr. Trypan blue staining was detected within 1–3 hr (white bars), whereas DNA fragmentation (dUTP staining, black bars) developed slowly and incompletely after metabolic inhibition. B,Top panel, Ca2+ did not increase when astrocytes were inhibited metabolically in the absence of extracellular Ca2+. Middle panel, Uncoupling was less pronounced when metabolic inhibition was not associated with [Ca2+]i increments. Bottom panel, The pattern of astrocytic death evoked by metabolic inhibition proceeded independently of concomitant [Ca2+]i changes. C,Top panel, Fifteen-minute exposure to the calcium ionophore lasalocid is associated with a surge in [Ca2+]i. Middle panel, The increase in [Ca2+]i is associated with a transient decrease in refill after photobleach. After normalization of [Ca2+]i to near resting levels, coupling slowly improved but declined again concomitant with a terminal increase in [Ca2+]i.Bottom panel, Ionophore exposure resulted in generalized death within 3 hr. These observations indicate that the permeability of astrocytic gap junctions is lowered, but not blocked, after lethal injury and that increments in Ca2+ partially are responsible for the decrease in coupling. *FRAP values compared with control values; p > 0.01. Data obtained 0–25, 25–50, 50–75, 75–100, 100–125, and 125–150 min after metabolic inhibition or ionophore exposure were pooled; two-way ANOVA was used to compare mean values, and Dunnett’s multiple range test was used to establish significant differences between control values and treatment values. §FRAP values compared with FRAP values 0–25 min after initiation of ionophore exposure [31 ± 5% (0–25 min),n = 25 vs 65 ± 8% (50–75 min),n = 12 and 51 ± 10% (75–100 min),n = 11; p > 0.01]. Thus, coupling increased significantly after ionophore exposure in astrocytes destined to die and was significantly higher 50–100 min after ionophore exposure than during the initial 0–25 min. The calcium tracings in the top panels plot data from representative cells. A total of 150 cells were analyzed in 15 individual experiments in A–C.
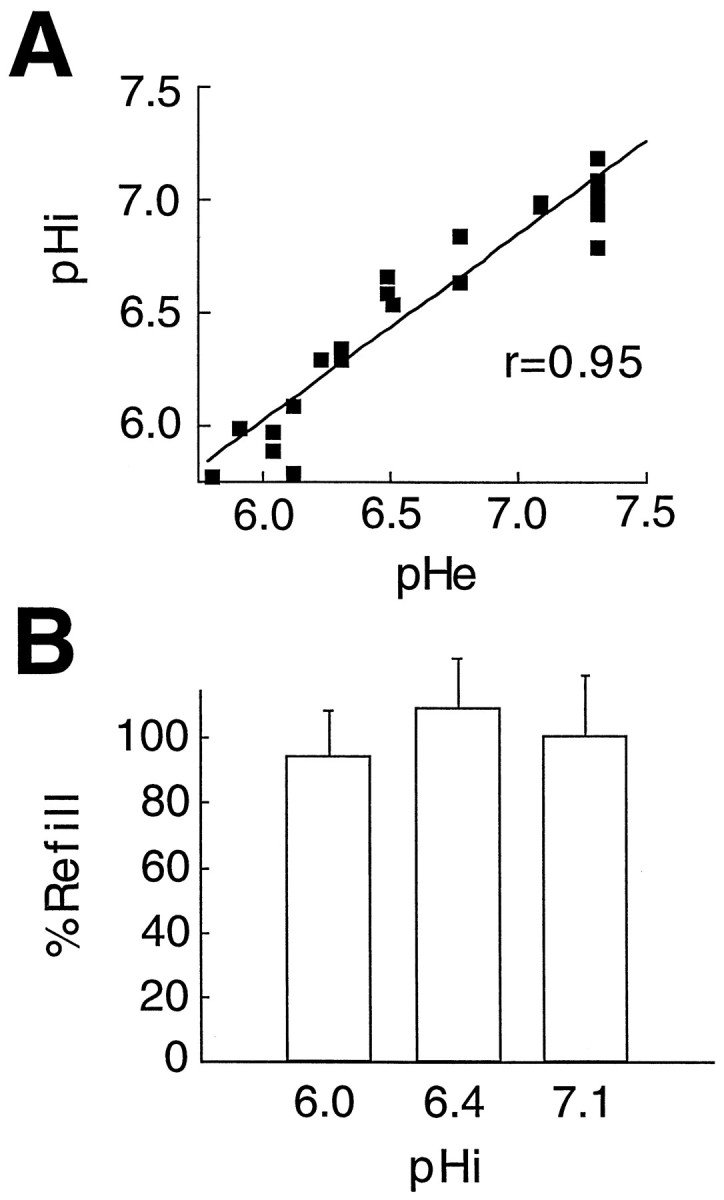
Acidosis does not modulate astrocytic couplingin vitro. A, To quantify coupling as a function of pHi, we first evaluated the relation of pHi and pHe in astrocytes loaded with the pH indicators BCECF and DCF. Exposure to an acidified lactic acid-containing solution resulted in a parallel reduction in pHi and pHe. Thus, pHi is a direct function of pHe in cultured astrocytes. B, Fluorescence recovery after photobleach was not altered significantly when pHi was lowered from 7.0 during control conditions to either pHi 6.4 or 6.0.
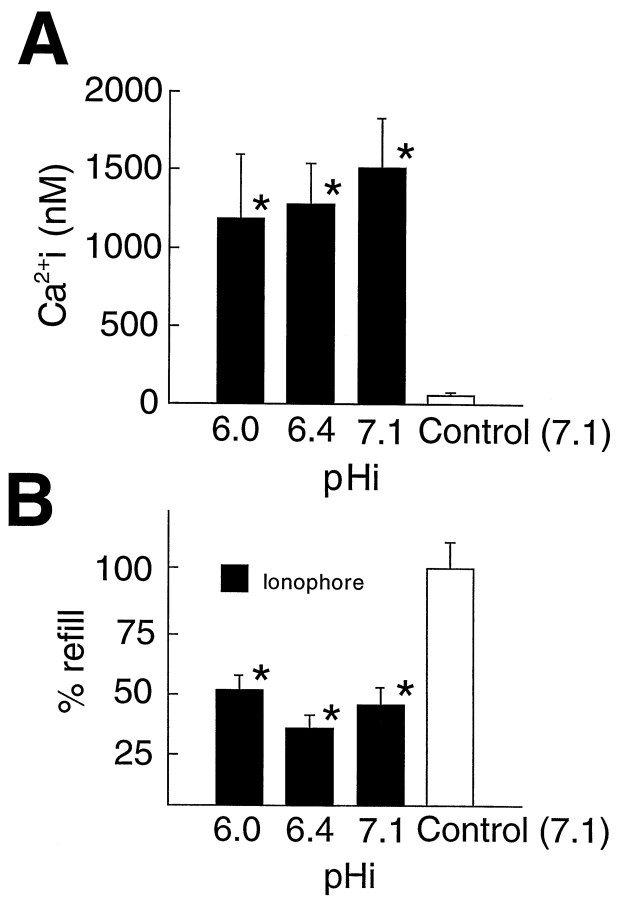
Acidosis does not potentiate the uncoupling effect of high [Ca2+]i.A, [Ca2+]i increments evoked by ionophore treatment were not affected significantly by concomitant acidosis. Controls were exposed to vehicle (0.4% DMF).B, Gap junction coupling detected by FRAP. The [Ca2+]i increase evoked by ionophore treatment reduced refill to 42 ± 6% of control values at pHi 7.0. Intracellular acidosis (pHi 6.0 and 6.4) did not decrease coupling further. Thus, intracellular acidosis does not potentiate the action of Ca2+ in reducing gap junction function. Two-way ANOVA was used to compare mean values, and Dunnett’s multiple range test was used to establish significant differences between treatment groups and controls. *Denotes significant difference at p < 0.01 relative to control. Error bars represent the mean ± SEM.
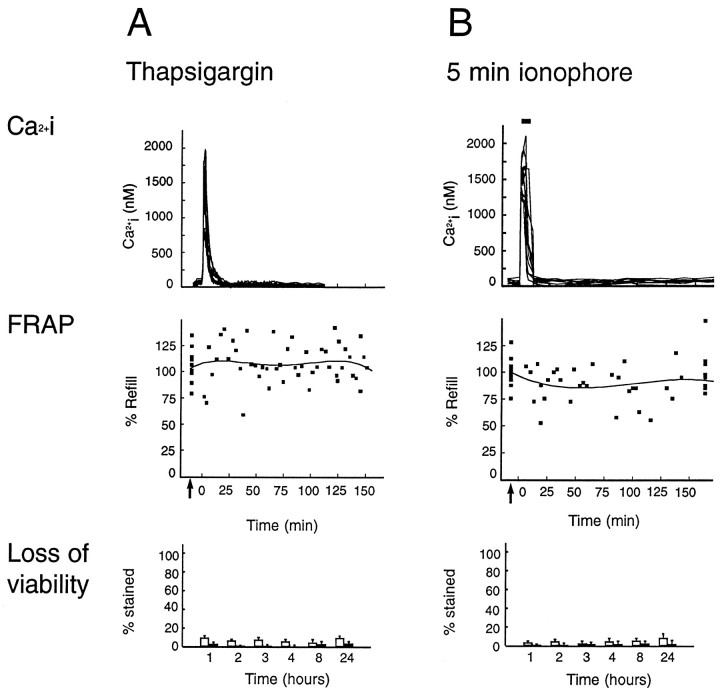
Nonlethal [Ca2+]i increments are not associated with a significant decrease in astrocytic coupling. Shown is a comparison of [Ca2+]i (top panels), gap junction coupling normalized against control values (middle panels), and loss of viability (bottom panels) during and after nonlethal [Ca2+]i increments. A,Top panel, A rapid increase in [Ca2+]i was evoked by thapsigargin (1 μm ). No sign of decrease in coupling was observed (middle panel). Refill after photobleach in separate experiments is mapped individually (squares) and the data fit by four-order regression. Bottom panel, The percentage of cell death after the same procedure. No loss of viability was observed by either trypan blue staining (white bars) or TUNEL stain (dUTP staining, black bars) at any time point. B, Top panel, [Ca2+]i increments during 5 min exposure to ionophore (lasalocid, 40 μm ). Middle panel, Uncoupling was not observed when ionophore exposure was not associated with the loss of viability. Bottom panel, No astrocytic death was evoked by short-term ionophore exposure. These observations indicate that nonlethal increments in Ca2+ do not significantly lower the permeability of astrocytic gap junctions.
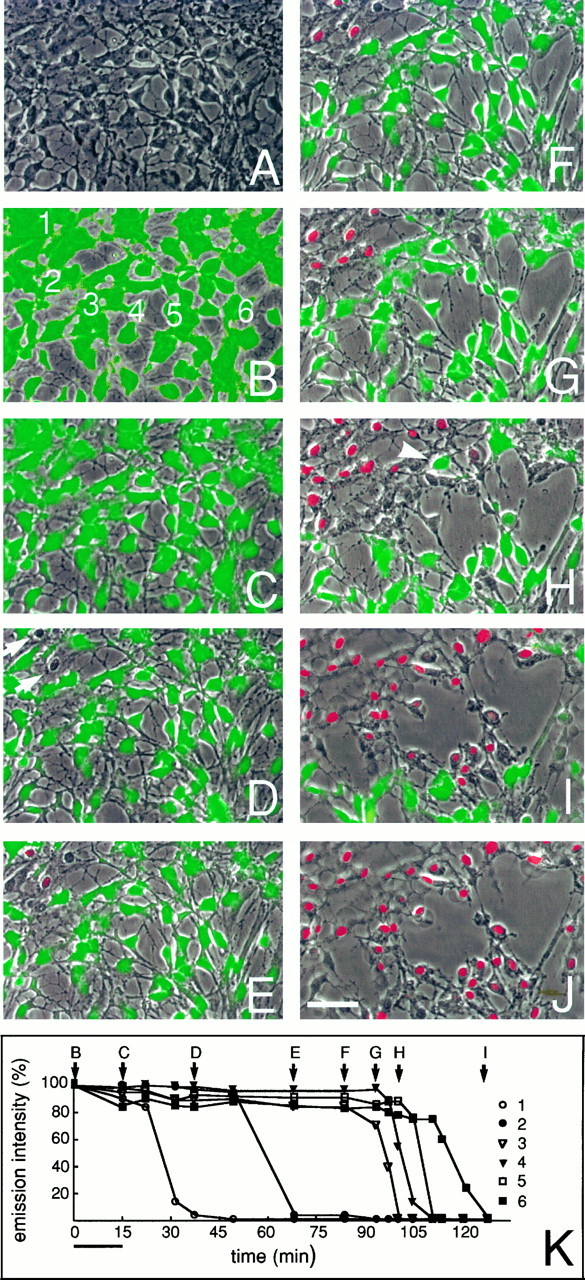
Astrocytic gap junctions close concurrently with the loss of membrane integrity. These photos depict the temporal and spatial pattern of cell death after exposure to ionophore. The culture was preloaded with CDCF, and the HBSS contained propidium iodide (2 μm ). A, Phase-contrast micrograph of a field shortly before exposure to the calcium ionophore.B, C, CDCF fluorescence digitally overlaid on the phase image of the same field before (B) and immediately after (C) ionophore exposure. All cells remained viable at this stage, but a general loss of CDCF fluorescence occurred during ionophore exposure. D, At 40 min after exposure, two cells have lost membrane integrity, and thereby CDCF fluorescence, but have not yet incorporated propidium iodide (arrows).E, At 70 min, injury has progressed to include four cells. Nuclear staining with propidium iodide (red) is now detectable in two cells. F–J, After 80–128 min, injury progressed with the recruitment of additional dying astrocytes. With further progression, cells began lifting from the substrate layer, leaving the plane of focus (top left corner). The gradual expansion of injury continued for the most part contiguously and moved across the field with a velocity of ∼3 μm/min. Note that CDCF fluorescence was retained until membrane integrity was lost, even in cells surrounded by dead cells (arrowhead).K, Relative emission intensity of CDCF in six labeled cells (B) during progression of injury. CDCF fluorescence is mapped as a function of time. Fluorescence remains relatively constant until membrane ruptures. Scale bar, 50 μm.
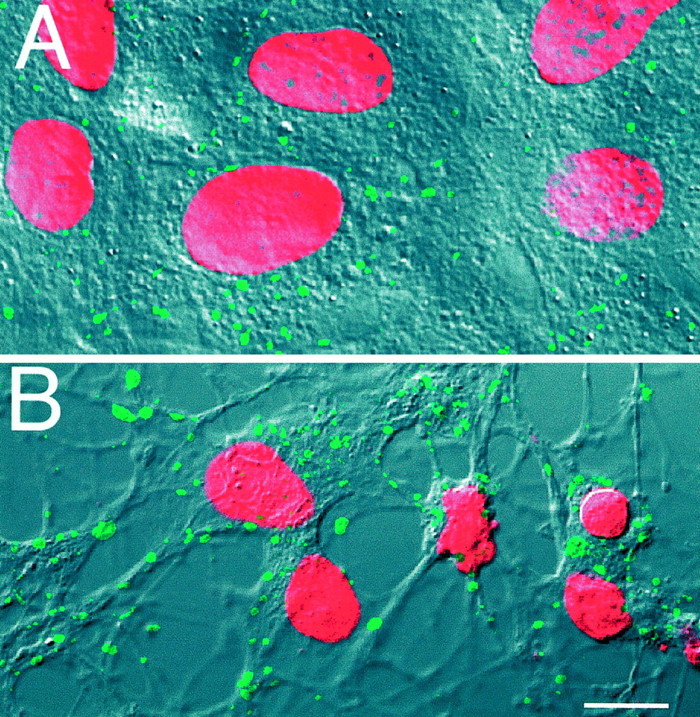
Loss of astrocytic viability is associated with the aggregation of Cx43 immunoreactive plaques. A, Cx43 immunoreactive plaques (fluorescein) are localized in the plasma membrane, with preference for sites of cell-to-cell contact. The culture was counterstained with propidium iodide (red) after fixation to quantify the number of plaques per nucleus. The fluorescence images were superimposed on DIC so that the distribution of gap junction plaques could be visualized. B, Two hours after metabolic inhibition, the number of Cx43 immunoreactive plaques had decreased concurrently with an increase in plaque size. Plaques remained in the plasma membrane. Note the decrease in nuclear size associated with the loss of viability.
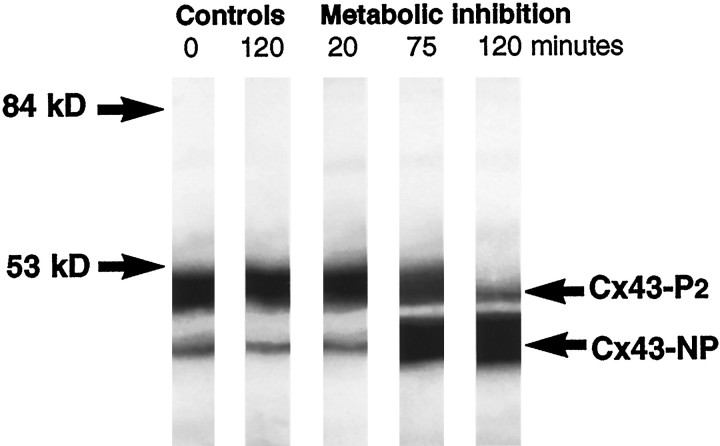
Western blot analysis of Cx43. A 20 min metabolic inhibition did not change the ratio between unphosphorylated Cx43 (Cx43-NP) and phosphorylated Cx43 (Cx43-P2), as compared with control. However, a partial dephosphorylation had taken place at 75 min and progressed by 120 min. Dephosphorylation parallels functional uncoupling (see Fig. 8A, middle panel), supporting the notion that Cx43-P2is required for the formation of functional channels. Two control samples, one collected directly and one incubated in HBSS for 2 hr, are shown. Prolonged incubation in HBSS was not associated with detectable changes in phosphorylation pattern.
References
-
- Anders JJ. Lactic acid inhibition of gap junctional intercellular communication in in vitro astrocytes as measured by fluorescence recovery after laser photobleaching. Glia. 1988;1:371–379. - PubMed
-
- Arellano G, Ramon F, Rivera A, Zampighi G. Calmodulin acts as an intermediary for the effects of calcium on gap junctions from crayfish lateral axons. J Membr Biol. 1988;101:119–131. - PubMed
-
- Astrup J, Symon L, Branston N, Lassen N. Cortical evoked potential and extracellular potassium and hydrogen at cortical levels of brain ischemia. Stroke. 1977;8:51–57. - PubMed
-
- Astrup J, Siesjo B, Symon L. Threshold in cerebral ischemia—the ischemic penumbra. Stroke. 1981;12:723–725. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous