

Cl- transport by cystic fibrosis transmembrane conductance regulator (CFTR) contributes to the inhibition of epithelial Na+ channels (ENaCs) in Xenopus oocytes co-expressing CFTR and ENaC
- PMID: 9518736
- PMCID: PMC2230925
- DOI: 10.1111/j.1469-7793.1998.825bp.x
Cl- transport by cystic fibrosis transmembrane conductance regulator (CFTR) contributes to the inhibition of epithelial Na+ channels (ENaCs) in Xenopus oocytes co-expressing CFTR and ENaC
Abstract
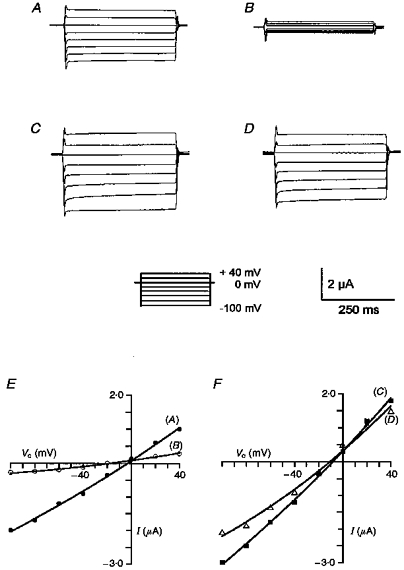
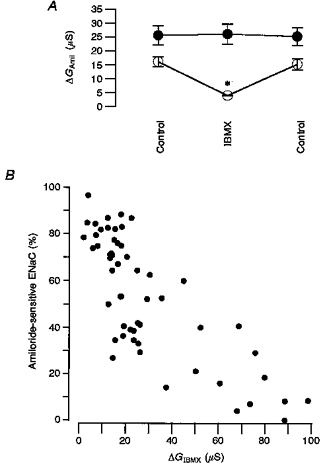
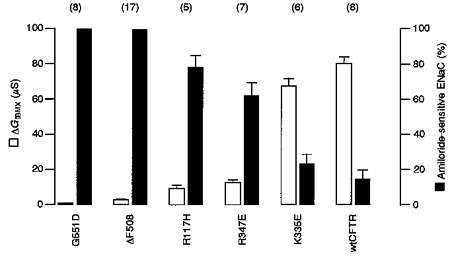
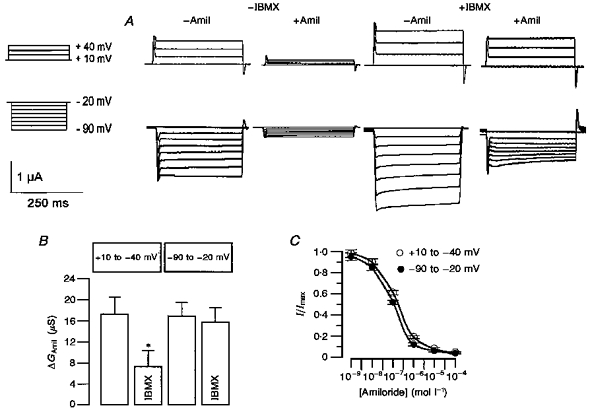
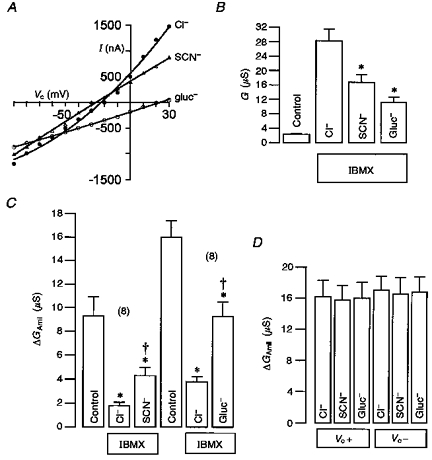
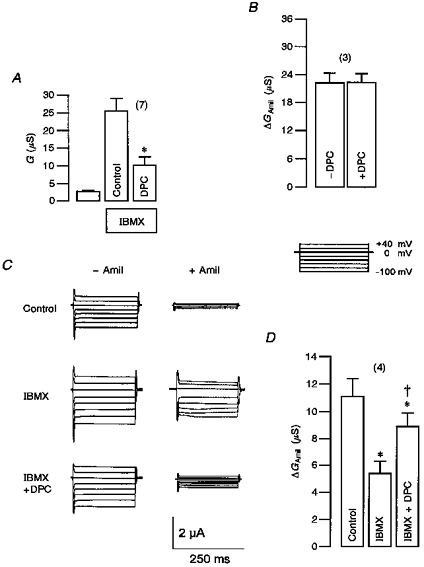
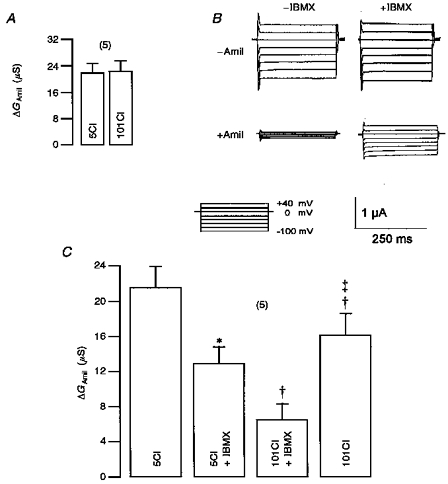
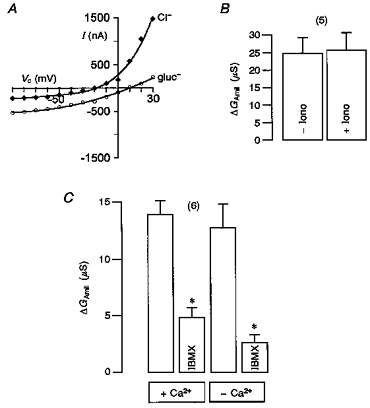
1. Epithelial Na+ channels (ENaCs) are inhibited by the cystic fibrosis transmembrane conductance regulator (CFTR) when CFTR is activated by protein kinase A. Since cAMP-dependent activation of CFTR Cl- conductance is defective in cystic fibrosis (CF), ENaC currents are not inhibited by CFTR. This could explain the enhanced Na+ conductance found in CF. In the present study, we examined possible mechanisms of interaction between CFTR and ENaC co-expressed in Xenopus oocytes. 2. The magnitude of CFTR Cl- currents activated by 3-isobutyl-1-methylxanthine (IBMX) in oocytes co-expressing either wild-type or mutant CFTR and ENaC determined the degree of downregulation of ENaC currents. 3. The ability of CFTR to inhibit ENaC currents was significantly reduced either when extracellular Cl- was replaced by poorly conductive anions, e.g. SCN- or gluconate, or when CFTR was inhibited by diphenylamine-carboxylate (DPC, 1 mmol l-1). 4. Downregulation of ENaC was more pronounced at positive when compared with negative clamp voltages. This suggests that outward currents, i.e. influx of Cl- through activated CFTR most effectively downregulated ENaC. 5. Activation of endogenous Ca2+-activated Cl- currents by 1 micromol l-1 ionomycin did not inhibit ENaC current. This suggests that inhibition of ENaC mediated by Cl- currents may be specific to CFTR. 6. The present findings indicate that downregulation of ENaC by CFTR is correlated to the ability of CFTR to conduct Cl-. The data have implications for how epithelia switch from NaCl absorption to NaCl secretion when CFTR is activated by secretagogues.
Figures
References
-
- Anderson M P, Gregory R J, Thompson S, Souza D W, Sucharita P, Mulligan R C, Smith A E, Welsh M J. Demonstration that CFTR is a chloride channel by alteration of its anion selectivity. Science. 1991;253:202–205. - PubMed
-
- Ballard S T, Fountain J D, Inglis S K, Corboz M R, Taylor A E. Chloride secretion across distal airway epithelium: relationship to submucosal gland distribution. American Journal of Physiology. 1995;268:L526–531. - PubMed
-
- Berdiev B K, Prat A G, Cantiello H F, Ausiello D A, Fuller C M, Jovov B, Benos D J, Ismailov I I. Regulation of epithelial sodium channels by short actin filaments. Journal of Biological Chemistry. 1996;271:17704–17710. 10.1074/jbc.271.30.17704. - DOI - PubMed
-
- Bradbury N A, Jilling T, Gabor B, Sorscher E J, Bridges R J, Kirk K L. Regulation of plasma membrane recycling by CFTR. Science. 1992;256:530–531. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous