

Structural analysis of the Spiroplasma virus, SpV4: implications for evolutionary variation to obtain host diversity among the Microviridae
- PMID: 9519405
- PMCID: PMC4167680
- DOI: 10.1016/s0969-2126(98)00016-1
Structural analysis of the Spiroplasma virus, SpV4: implications for evolutionary variation to obtain host diversity among the Microviridae
Abstract
Background: Spiroplasma virus, SpV4, is a small, non-enveloped virus that infects the helical mollicute Spiroplasma melliferum. SpV4 exhibits several similarities to the Chlamydia phage, Chp1, and the Coliphages alpha 3, phi K, G4 and phi X174. All of these viruses are members of the Microviridae. These viruses have isometric capsids with T = 1 icosahedral symmetry, cause lytic infections and are the only icosahedral phages that contain single-stranded circular DNA genomes. The aim of this comparative study on these phages was to understand the role of their capsid proteins during host receptor recognition.
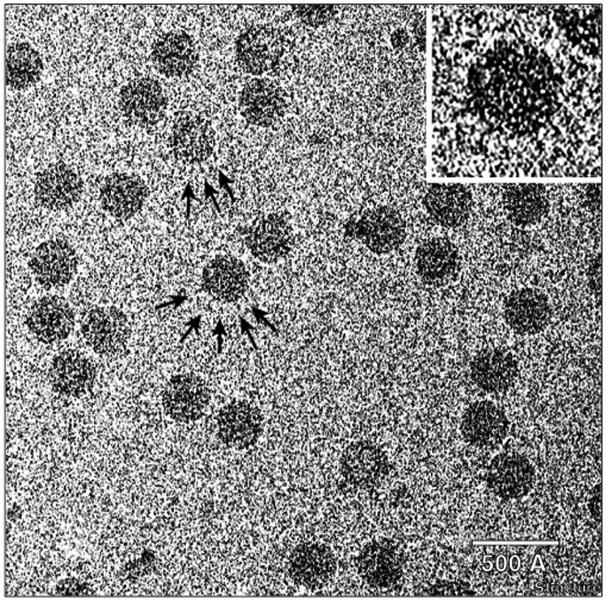
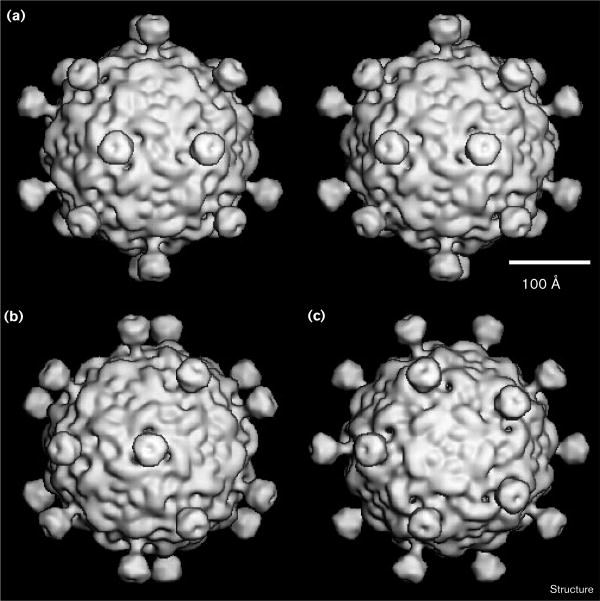
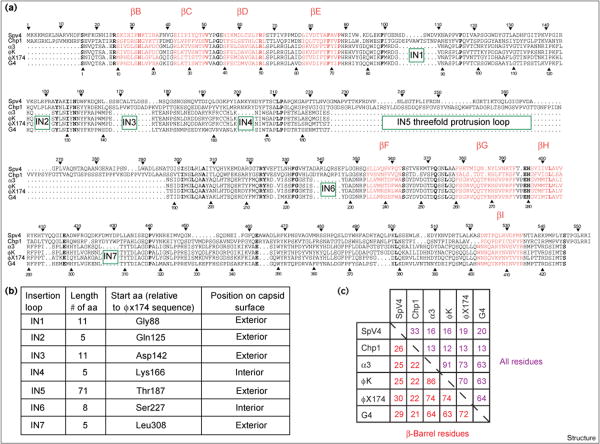
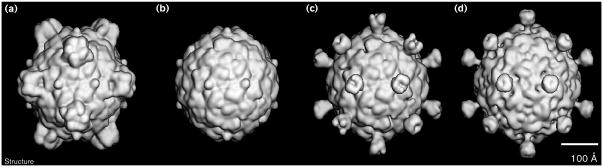
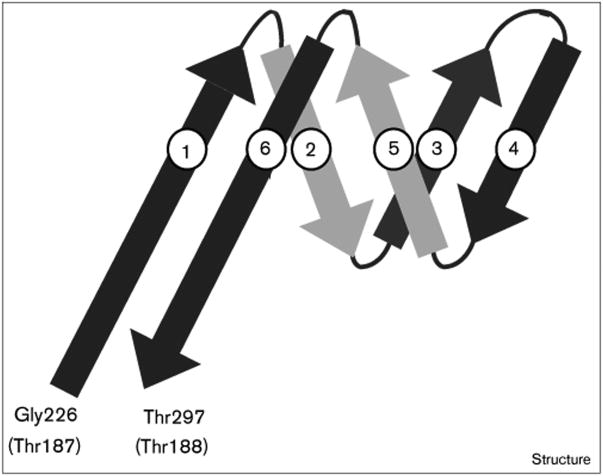
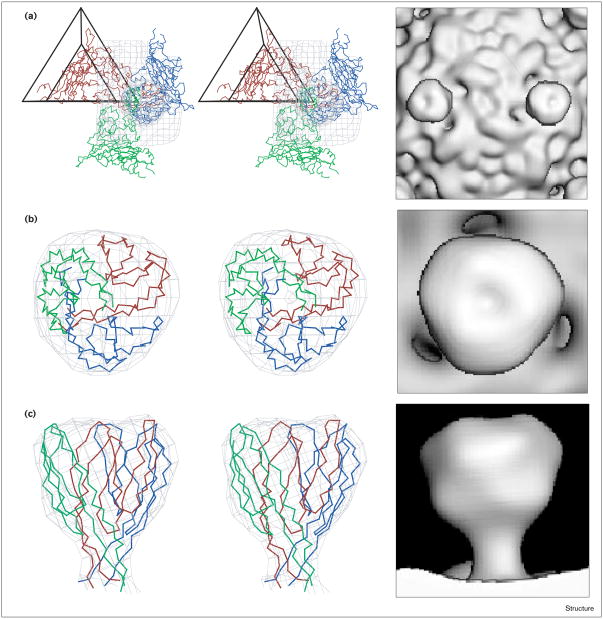
Results: The three-dimensional structure of SpV4 was determined to 27 A resolution from images of frozen-hydrated particles. Cryo-electron microscopy (cryo-EM) revealed 20, approximately 54 A long, 'mushroom-like' protrusions on the surface of the capsid. Each protrusion comprises a trimeric structure that extends radially along the threefold icosahedral axes of the capsid. A 71 amino acid portion of VP1 (the SpV4 capsid protein) was shown, by structural alignment with the atomic structure of the F capsid protein of phi X174, to represent an insertion sequence between the E and F strands of the eight-stranded antiparallel beta-barrel. Secondary structure prediction of this insertion sequence provided the basis for a probable structural motif, consisting of a six-stranded antiparallel beta sheet connected by small turns. Three such motifs form the rigid stable trimeric structures (mushroom-like protrusions) at the threefold axes, with hydrophobic depressions at their distal surface.
Conclusions: Sequence alignment and structural analysis indicate that distinct genera of the Microviridae might have evolved from a common primordial ancestor, with capsid surface variations, such as the SpV4 protrusions, resulting from gene fusion events that have enabled diverse host ranges. The hydrophobic nature of the cavity at the distal surface of the SpV4 protrusions suggests that this region may function as the receptor-recognition site during host infection.
Figures




References
-
- Ricard B, Garnier M, Bove JM. Characterization of SPV3 from spiroplasmas and discovery of a new spiroplasma virus (SPV4) Rev Infect Dis. 1982;4:S275.
-
- Renaudin J, Pascarel MC, Garnier M, Carle P, Bove JM. Characterization of spiroplasma virus group 4 (SPV4) Isr J Med Sci. 1984;20:797–799. - PubMed
-
- Renaudin J, Pascarel MC, Garnier M, Junca-Carle P, Bove JM. SpV4, a new spiroplasma virus with circular, single stranded DNA. Ann Virol. 1984;135E:343–361.
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
