

A tripotential glial precursor cell is present in the developing spinal cord
- PMID: 9520481
- PMCID: PMC19951
- DOI: 10.1073/pnas.95.7.3996
A tripotential glial precursor cell is present in the developing spinal cord
Abstract
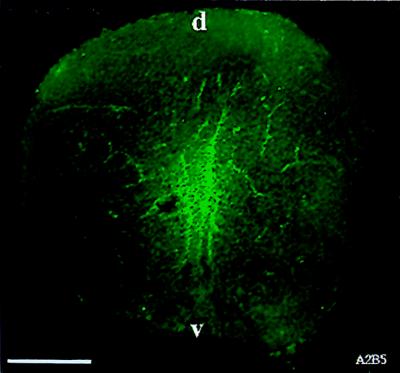
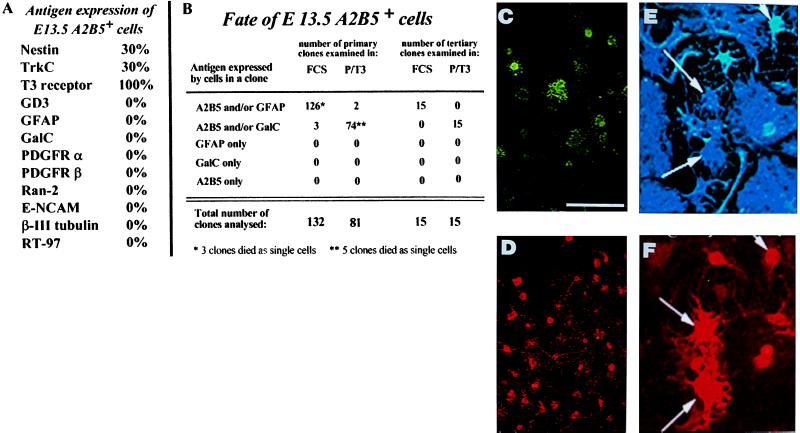
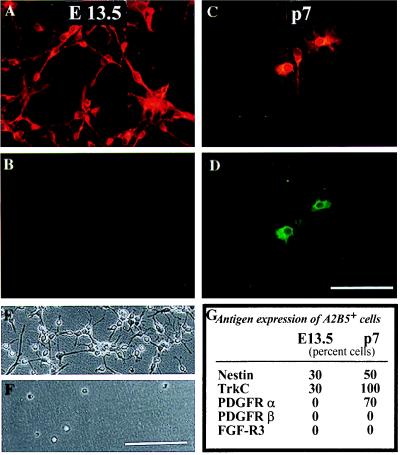
We have isolated a tripotential glial precursor cell population from spinal cords of E13.5 rats. In vitro, these A2B5+E-NCAM- glial-restricted precursor (GRP) cells can undergo extensive self-renewal, and can differentiate into oligodendrocytes and two distinct astrocyte populations, but do not differentiate into neurons. The differentiation potential of GRP cells is retained through at least three cycles of expansion and recloning. Unlike oligodendrocyte-type 2 astrocyte progenitor cells, freshly isolated GRP cells do not respond to platelet-derived growth factor as a mitogen or survival factor, nor do GRP cells differentiate into oligodendrocytes--or even survive--when plated in mitogen-free chemically defined medium. Exposure to fetal calf serum induces GRP cells to differentiate into A2B5- fibroblast-like astrocytes, whereas growth in the presence of basic fibroblast growth factor and ciliary neurotrophic factor induces the generation of A2B5+ process-bearing astrocytes. The early appearance of GRP cells during spinal cord development suggests that they may represent the earliest GRP cell population.
Figures




References
-
- Price J, Williams B, Grove E. Brain Pathol. 1992;2:23–29. - PubMed
-
- Sanes J R. Trends Neurosci. 1989;12:21–28. - PubMed
-
- Goldman J E. J Neuro-Oncol. 1995;24:61–64. - PubMed
-
- Noble M, Gutoswki N, Bevan K, Engel U, Linskey M, Urenjak J, Bhakoo K, Williams S. Glia. 1995;15:222–230. - PubMed
-
- Raff M C. Science. 1989;243:1450–1455. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
