

Inhibition of hepatitis B virus replication during adenovirus and cytomegalovirus infections in transgenic mice
- PMID: 9525579
- PMCID: PMC109701
- DOI: 10.1128/JVI.72.4.2630-2637.1998
Inhibition of hepatitis B virus replication during adenovirus and cytomegalovirus infections in transgenic mice
Abstract
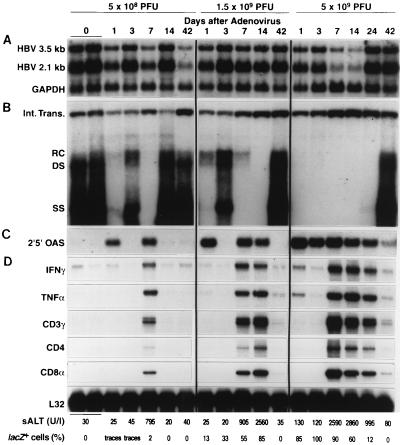
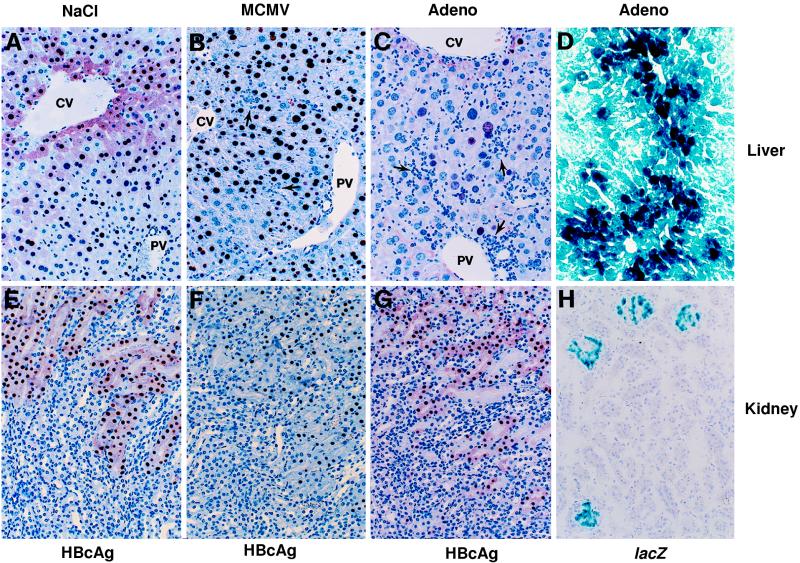
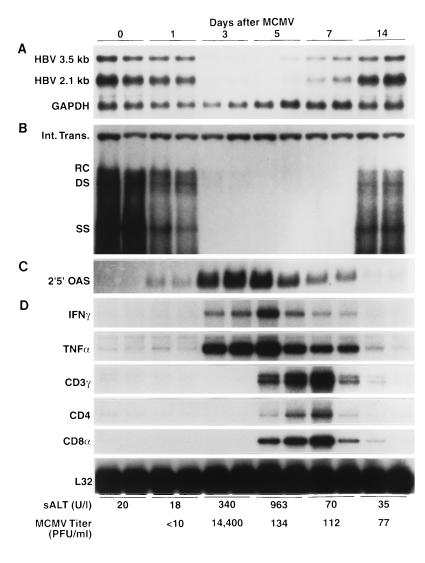
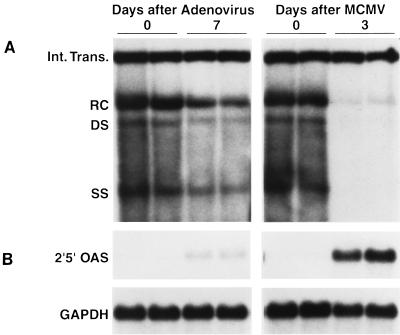
We have previously demonstrated that hepatitis B virus (HBV) replication and gene expression are abolished in the livers of HBV transgenic mice by cytotoxic T lymphocytes (CTLs) and during lymphocytic choriomeningitis virus (LCMV) infection, stimuli that trigger the production of alpha/beta interferon, gamma interferon, and tumor necrosis factor alpha in the liver. We now report that hepatic HBV replication and gene expression are inhibited by the local induction of these cytokines during adenovirus- and murine cytomegalovirus (MCMV)-induced hepatitis. Further, we show that MCMV also blocks HBV replication and gene expression in the proximal convoluted tubules of the kidney by causing interstitial nephritis and inducing the same cytokines in the renal parenchyma. These results suggest that inflammatory cytokines probably contribute to viral clearance during acute viral hepatitis in humans, and they imply that induction of these cytokines in the liver and other infected tissues of chronically infected patients might have therapeutic value.
Figures
References
-
- Bancroft G J, Shellam G R, Chalmer J E. Genetic influences on the augmentation of natural killer (NK) cells during murine cytomegalovirus infection: correlation with patterns of resistance. J Immunol. 1981;126:988–994. - PubMed
-
- Bouley D M, Kanangat S, Wire W, Rouse B T. Characterization of herpes simplex virus type-1 infection and herpetic stromal keratitis development in IFN-γ knockout mice. J Immunol. 1995;155:3964–3971. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
