

Cleavage of the feline calicivirus capsid precursor is mediated by a virus-encoded proteinase
- PMID: 9525628
- PMCID: PMC109753
- DOI: 10.1128/JVI.72.4.3051-3059.1998
Cleavage of the feline calicivirus capsid precursor is mediated by a virus-encoded proteinase
Abstract
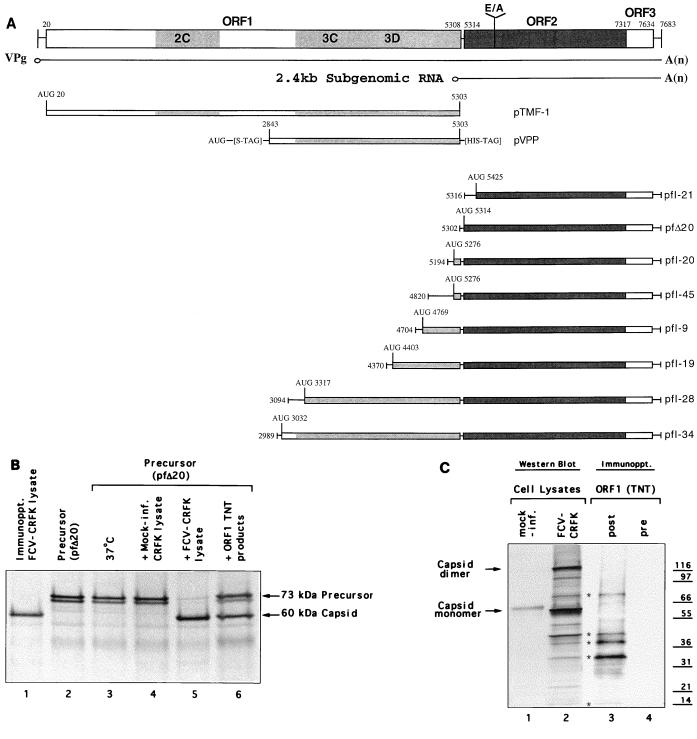
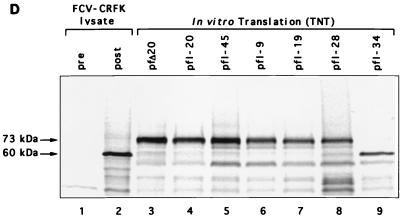
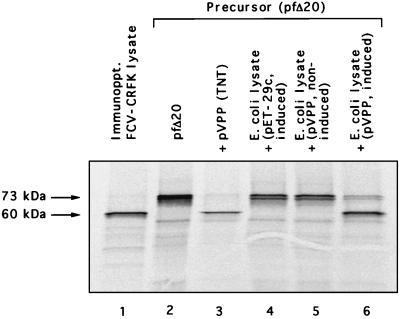
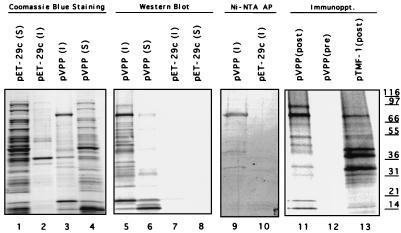
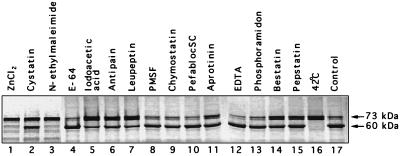
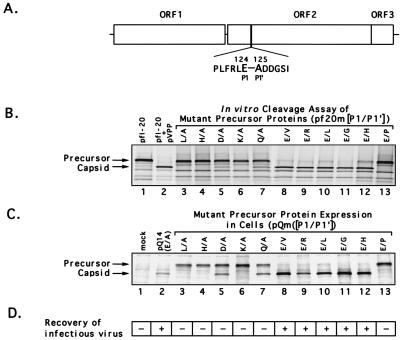
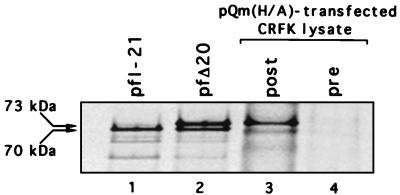
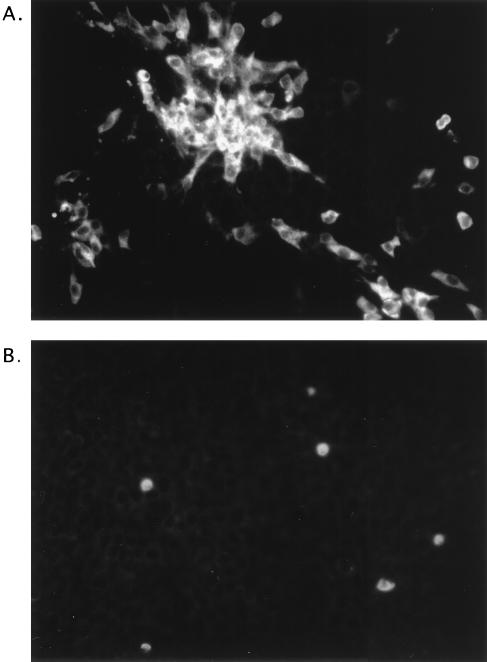
Feline calicivirus (FCV), a member of the Caliciviridae, produces its major structural protein as a precursor polyprotein from a subgenomic-sized mRNA. In this study, we show that the proteinase responsible for processing this precursor into the mature capsid protein is encoded by the viral genome at the 3'-terminal portion of open reading frame 1 (ORF1). Protein expression studies of either the entire or partial ORF1 indicate that the proteinase is active when expressed either in in vitro translation or in bacterial cells. Site-directed mutagenesis was used to characterize the proteinase Glu-Ala cleavage site in the capsid precursor, utilizing an in vitro cleavage assay in which mutant precursor proteins translated from cDNA clones were used as substrates for trans cleavage by the proteinase. In general, amino acid substitutions in the P1 position (Glu) of the cleavage site were less well tolerated by the proteinase than those in the P1' position (Ala). The precursor cleavage site mutations were introduced into an infectious cDNA clone of the FCV genome, and transfection of RNA derived from these clones into feline kidney cells showed that efficient cleavage of the capsid precursor by the virus-encoded proteinase is a critical determinant in the growth of the virus.
Figures
References
-
- Allaire M, Chernaia M M, Malcolm B A, James M N G. Picornaviral 3C cysteine proteinases have a fold similar to chymotrypsin-like serine proteinases. Nature. 1994;369:72–76. - PubMed
-
- Black D N, Burroughs J N, Harris T J R, Brown F. The structure and replication of calicivirus RNA. Nature. 1978;274:614–615. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
