

SWI-SNF complex participation in transcriptional activation at a step subsequent to activator binding
- PMID: 9528749
- PMCID: PMC121407
- DOI: 10.1128/MCB.18.4.1774
SWI-SNF complex participation in transcriptional activation at a step subsequent to activator binding
Abstract
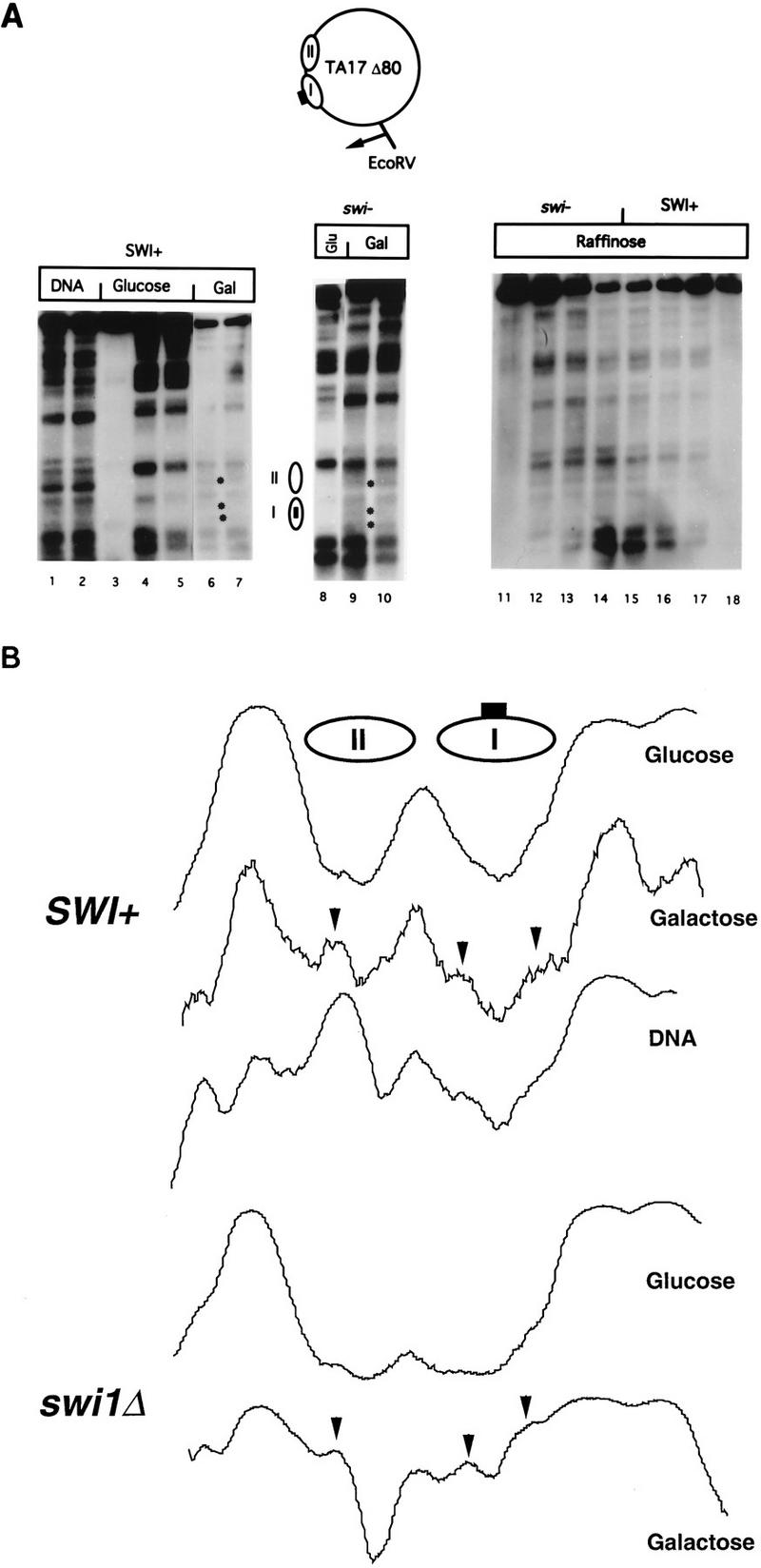
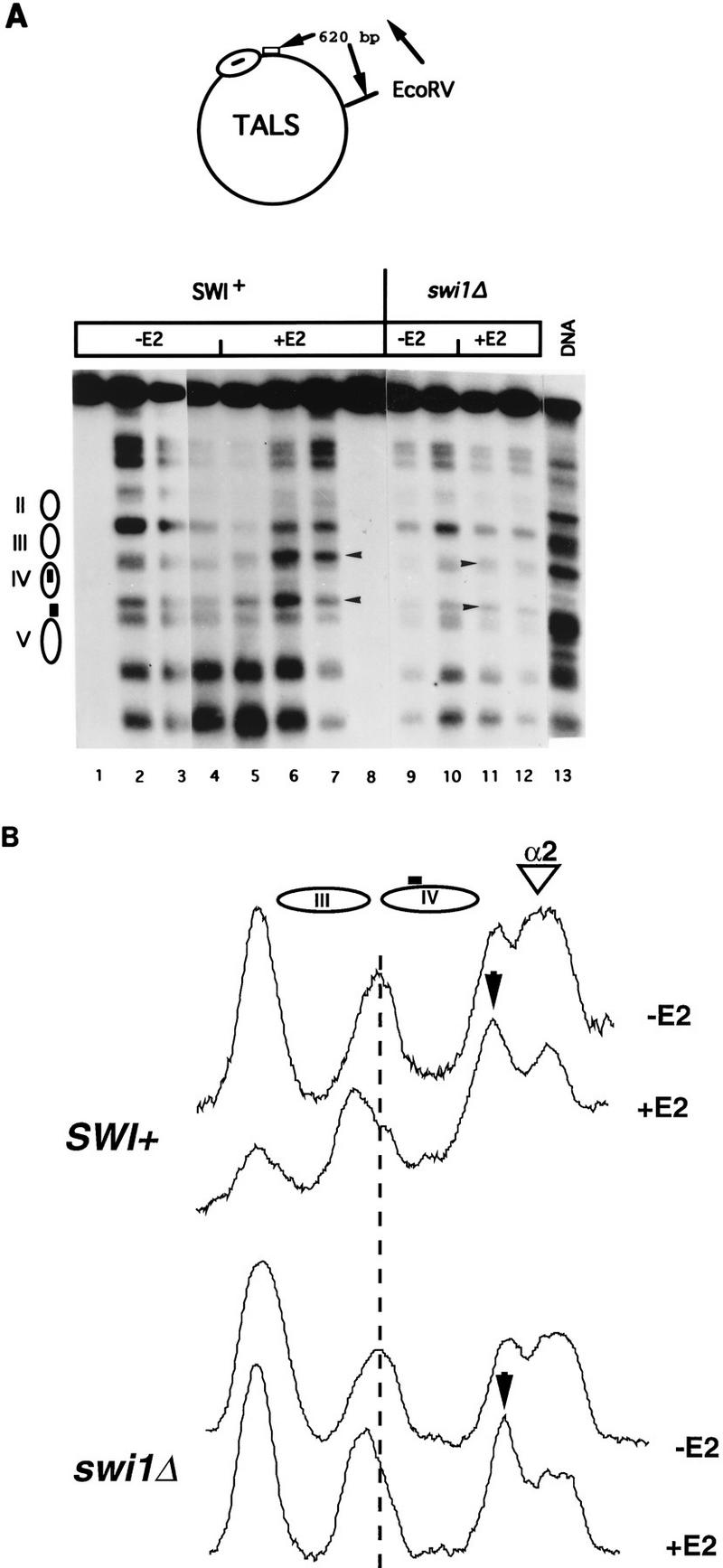
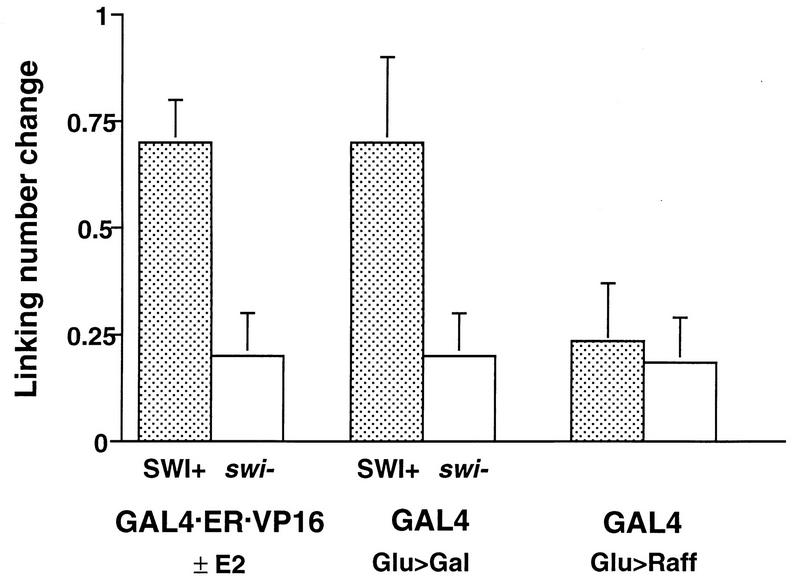
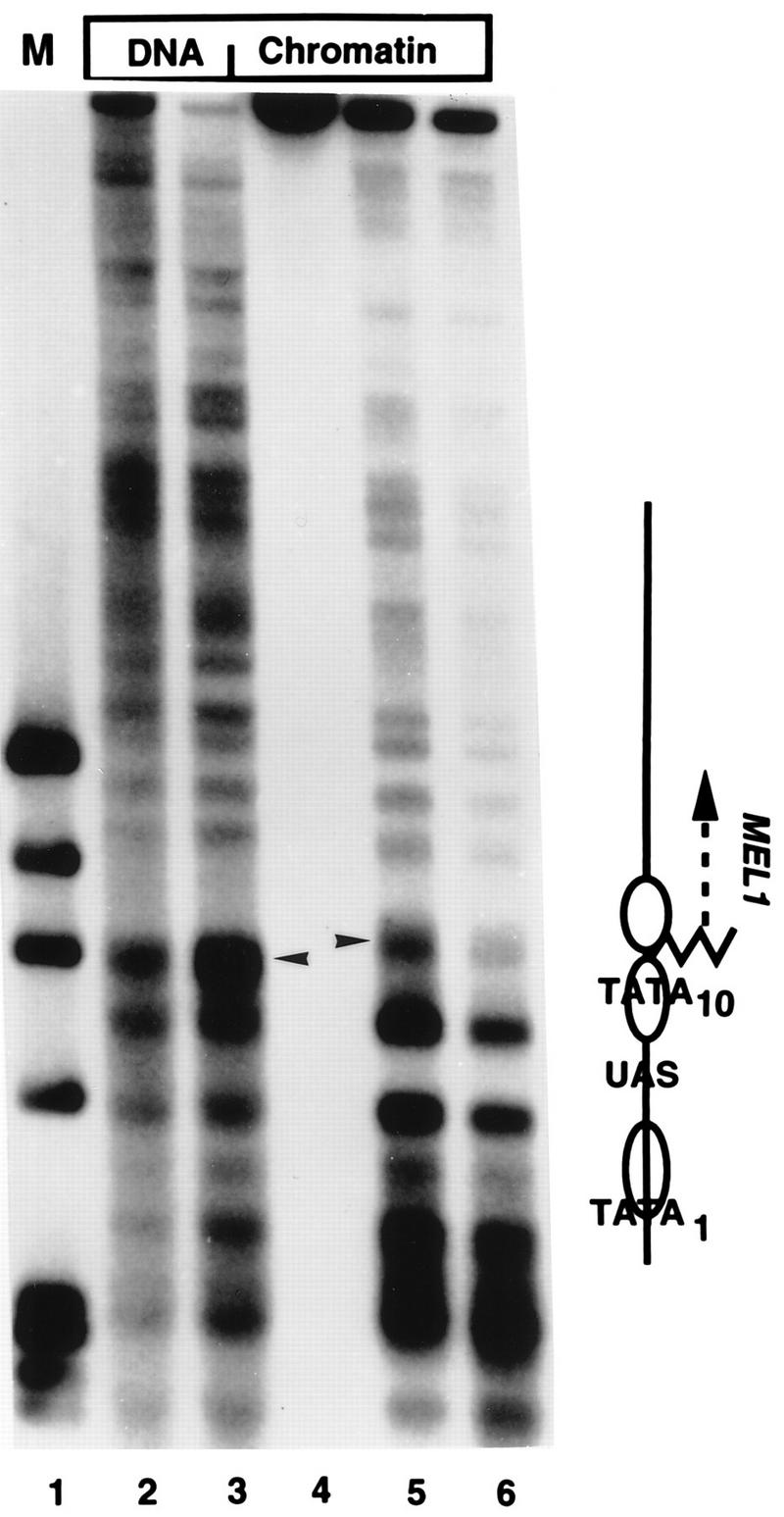
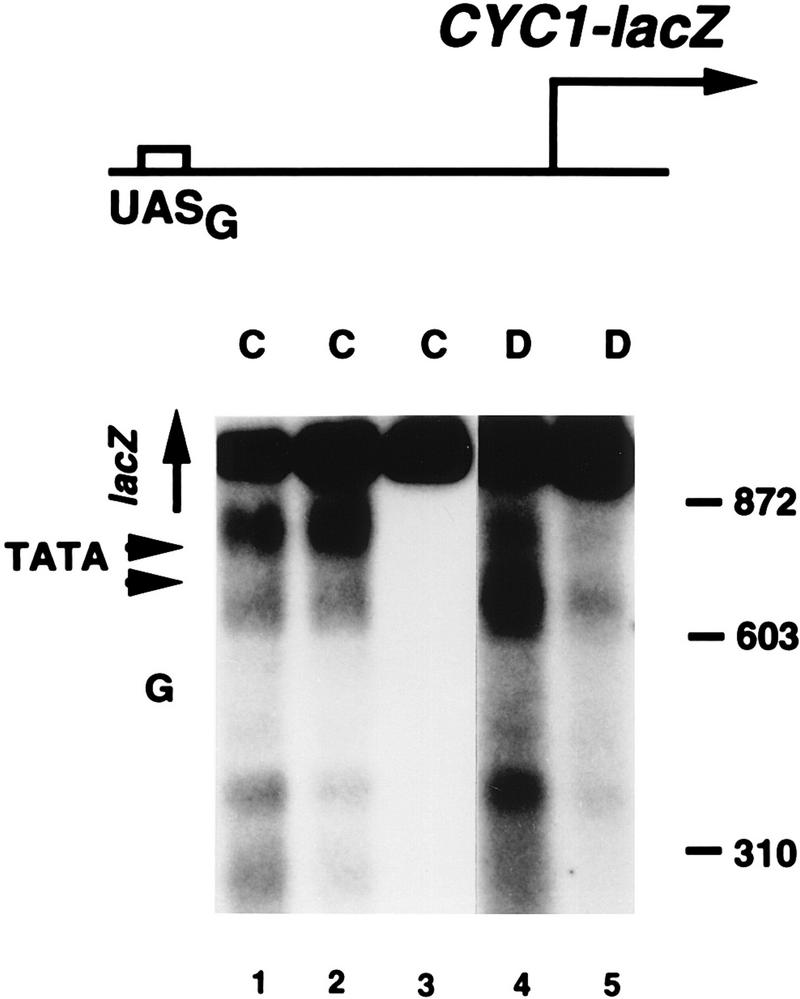
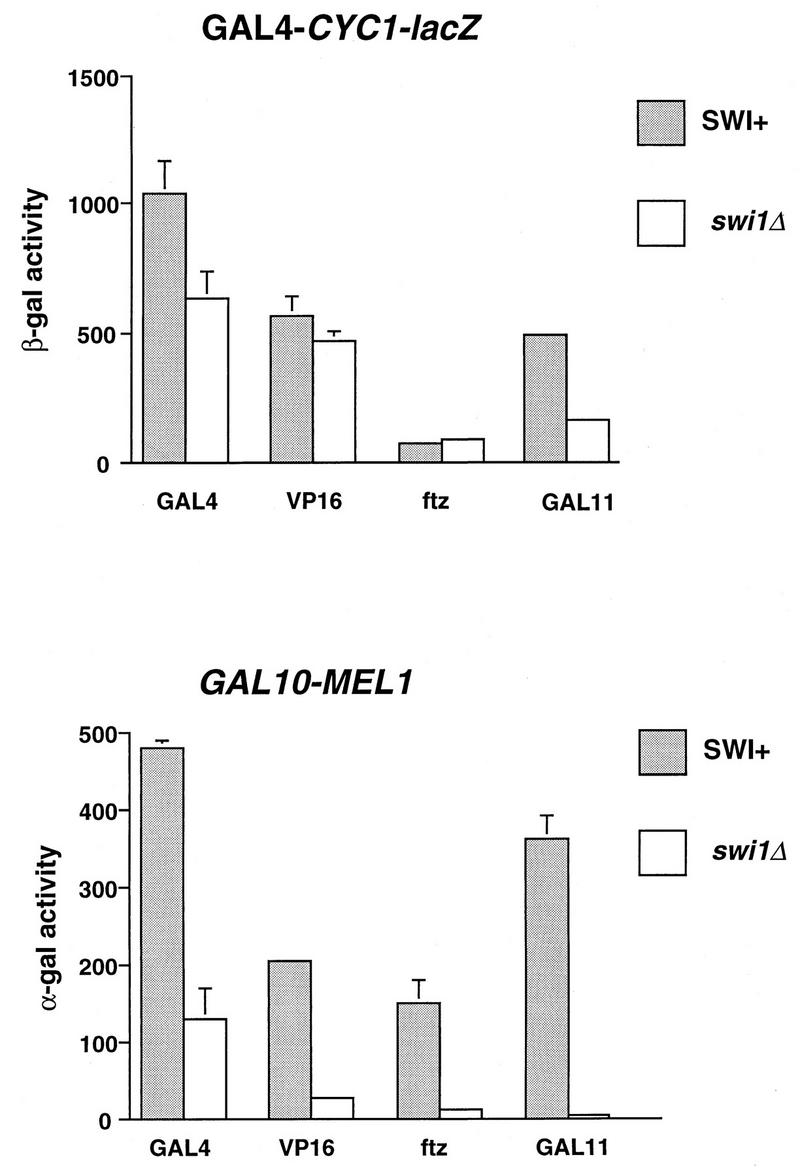
The SWI-SNF complex in yeast and related complexes in higher eukaryotes have been implicated in assisting gene activation by overcoming the repressive effects of chromatin. We show that the ability of the transcriptional activator GAL4 to bind to a site in a positioned nucleosome is not appreciably impaired in swi mutant yeast cells. However, chromatin remodeling that depends on a transcriptional activation domain shows a considerable, although not complete, SWI-SNF dependence, suggesting that the SWI-SNF complex exerts its major effect at a step subsequent to activator binding. We tested this idea further by comparing the SWI-SNF dependence of a reporter gene based on the GAL10 promoter, which has an accessible upstream activating sequence and a nucleosomal TATA element, with that of a CYC1-lacZ reporter, which has a relatively accessible TATA element. We found that the GAL10-based reporter gene showed a much stronger SWI-SNF dependence than did the CYC1-lacZ reporter with several different activators. Remarkably, transcription of the GAL10-based reporter by a GAL4-GAL11 fusion protein showed a nearly complete requirement for the SWI-SNF complex, strongly suggesting that SWI-SNF is needed to allow access of TFIID or the RNA polymerase II holoenzyme. Taken together, our results demonstrate that chromatin remodeling in vivo can occur by both SWI-SNF-dependent and -independent avenues and suggest that the SWI-SNF complex exerts its major effect in transcriptional activation at a step subsequent to transcriptional activator-promoter recognition.
Figures
References
-
- Barberis A J, Pearlberg J, Simkovich N, Farrell S, Reinagel P, Bamdad C, Sigal G, Ptashne M. Contact with a component of the polymerase II holoenzyme suffices for gene activation. Cell. 1995;81:359–368. - PubMed
-
- Brown S A, Imbalzano A N, Kingston R E. Activator-dependent regulation of transcriptional pausing on nucleosomal templates. Genes Dev. 1996;10:1479–1490. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous