

Rex-1, a gene encoding a transcription factor expressed in the early embryo, is regulated via Oct-3/4 and Oct-6 binding to an octamer site and a novel protein, Rox-1, binding to an adjacent site
- PMID: 9528758
- PMCID: PMC121416
- DOI: 10.1128/MCB.18.4.1866
Rex-1, a gene encoding a transcription factor expressed in the early embryo, is regulated via Oct-3/4 and Oct-6 binding to an octamer site and a novel protein, Rox-1, binding to an adjacent site
Abstract
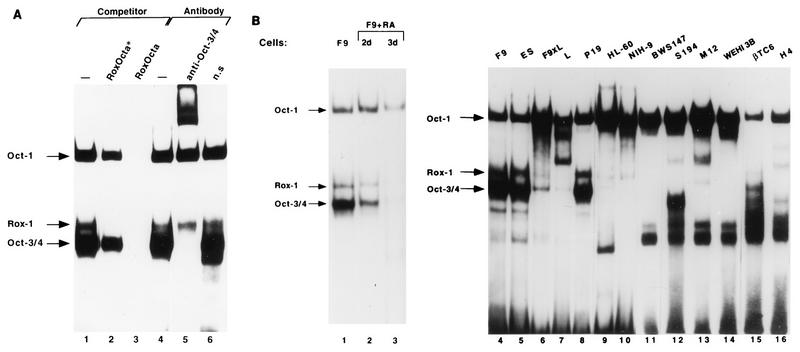
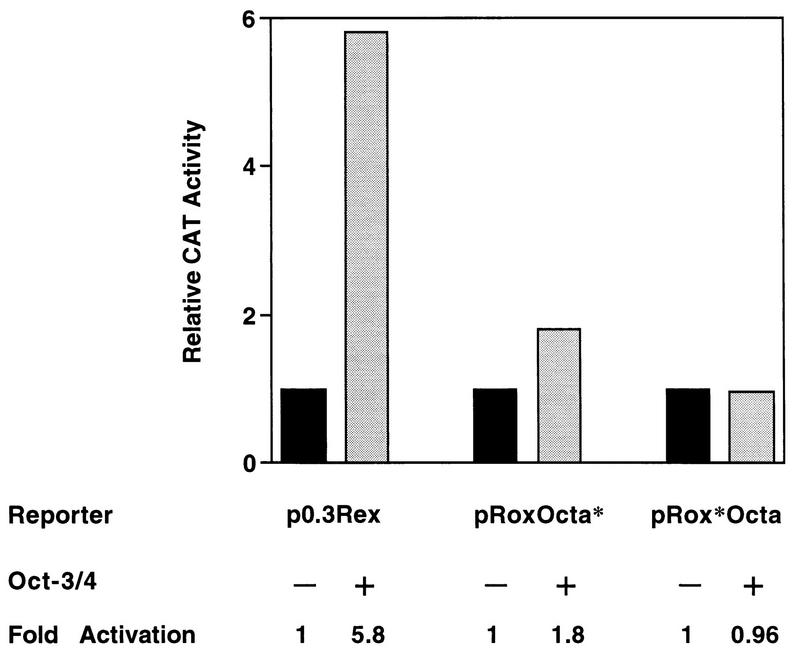
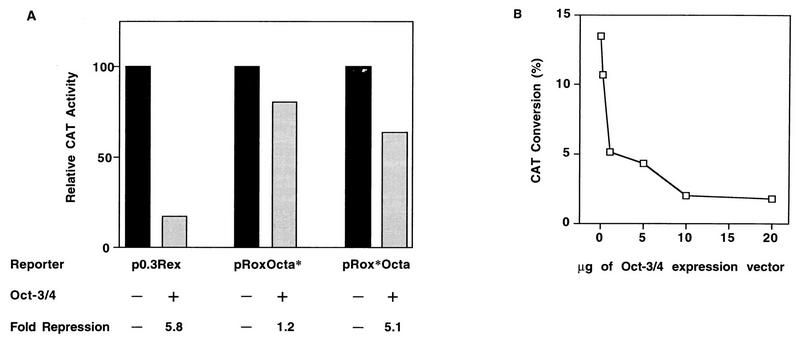
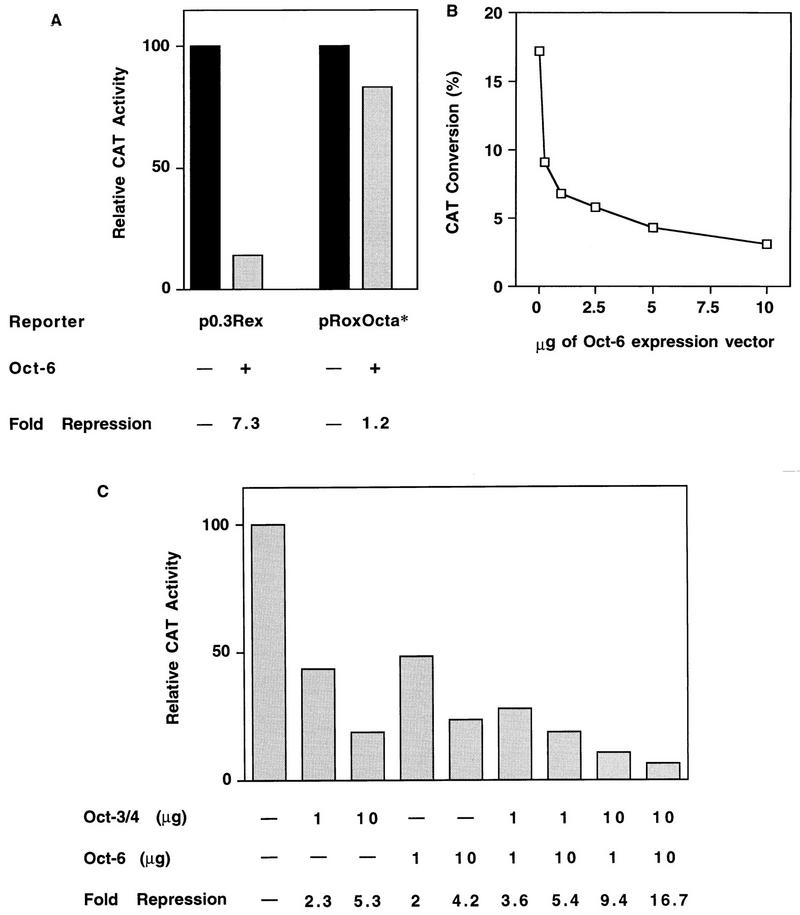
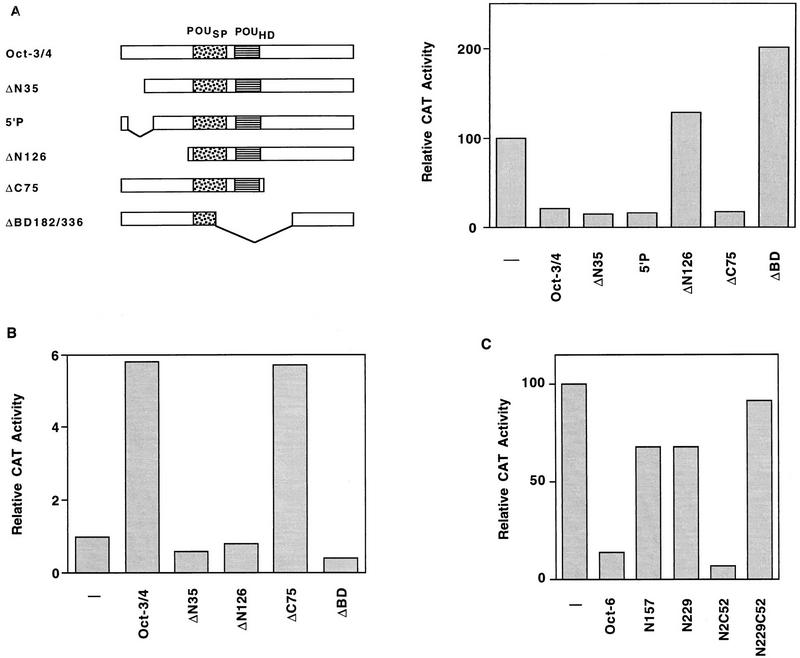
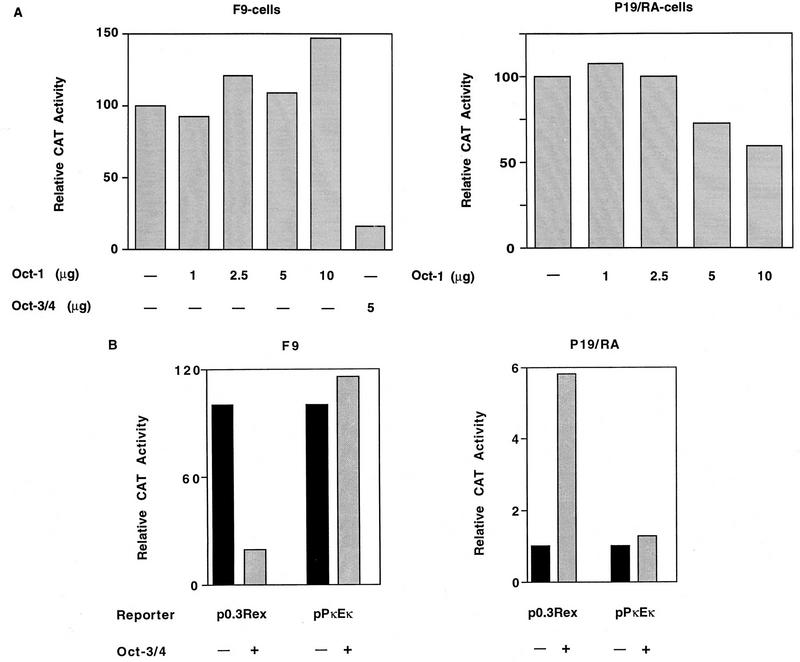
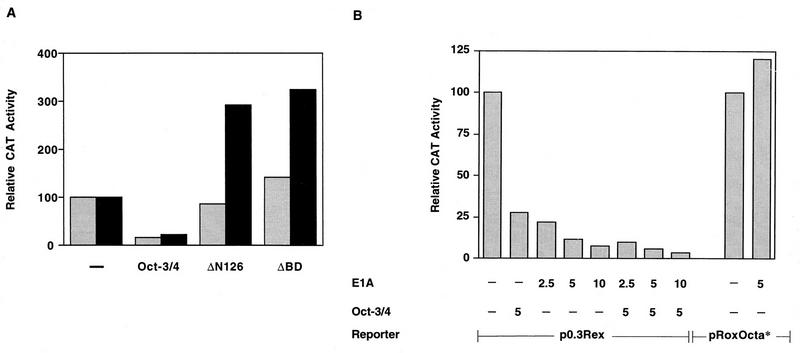
The Rex-1 (Zfp-42) gene, which encodes an acidic zinc finger protein, is expressed at high levels in embryonic stem (ES) and F9 teratocarcinoma cells. Prior analysis identified an octamer motif in the Rex-1 promoter which is required for promoter activity in undifferentiated F9 cells and is involved in retinoic acid (RA)-associated reduction in expression. We show here that the Oct-3/4 transcription factor, but not Oct-1, can either activate or repress the Rex-1 promoter, depending on the cellular environment. Rex-1 repression is enhanced by E1A. The protein domain required for Oct-3/4 activation was mapped to amino acids 1 to 35, whereas the domain required for Oct-3/4 repression was mapped to amino acids 61 to 126, suggesting that the molecular mechanisms underlying transcriptional activation and repression differ. Like Oct-3/4, Oct-6 can also lower the expression of the Rex-1 promoter via the octamer site, and the amino-terminal portion of Oct-6 mediates this repression. In addition to the octamer motif, a novel positive regulatory element, located immediately 5' of the octamer motif, was identified in the Rex-1 promoter. Mutations in this element greatly reduce Rex-1 promoter activity in F9 cells. High levels of a binding protein(s), designated Rox-1, recognize this novel DNA element in F9 cells, and this binding activity is reduced following RA treatment. Taken together, these results indicate that the Rex-1 promoter is regulated by specific octamer family members in early embryonic cells and that a novel element also contributes to Rex-1 expression.
Figures



References
-
- Ben-Shushan E, Pikarsky E, Klar A, Bergman Y. Extinction of Oct-3/4 gene expression in embryonal carcinoma × fibroblast somatic cell hybrids is accompanied by changes in the methylation status, chromatin structure, and transcriptional activity of the Oct-3/4 upstream region. Mol Cell Biol. 1993;13:891–901. - PMC - PubMed
-
- Bushmeyer S, Park K, Atchison M L. Characterization of functional domains within the multifunctional transcription factor, YY1. J Biol Chem. 1995;270:30213–30220. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous