

Yeast 18S rRNA dimethylase Dim1p: a quality control mechanism in ribosome synthesis?
- PMID: 9528805
- PMCID: PMC121492
- DOI: 10.1128/MCB.18.4.2360
Yeast 18S rRNA dimethylase Dim1p: a quality control mechanism in ribosome synthesis?
Abstract
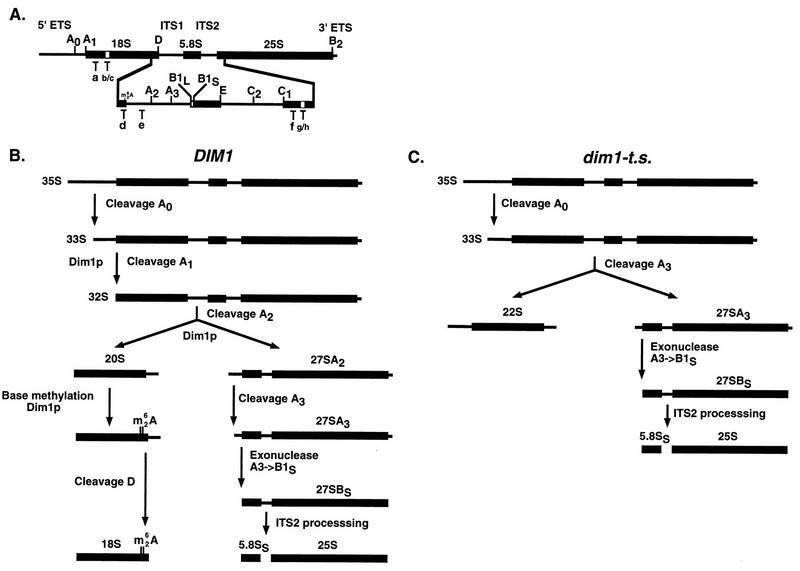
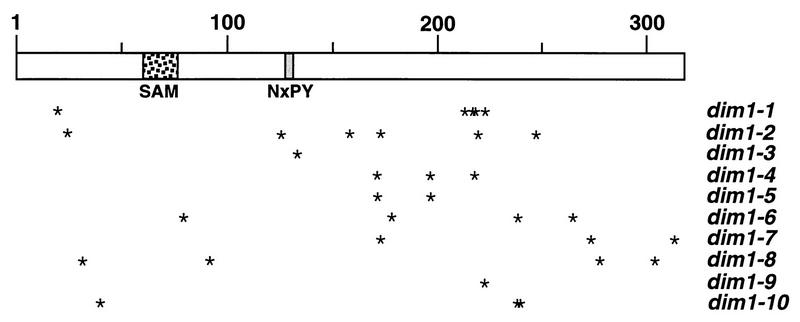
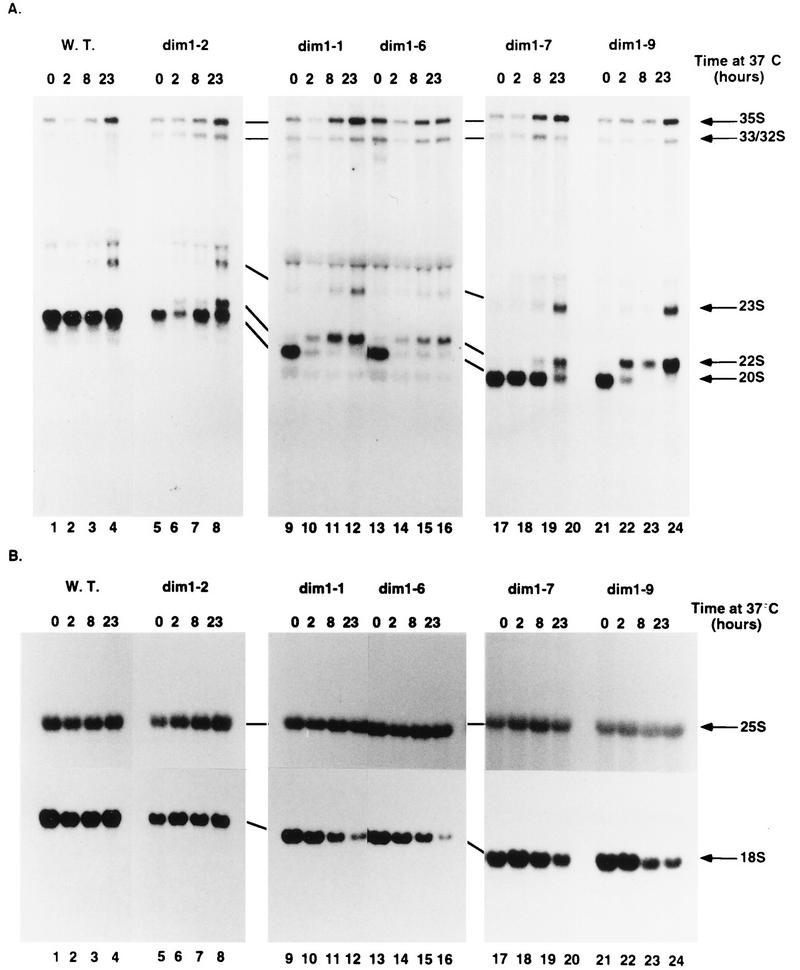
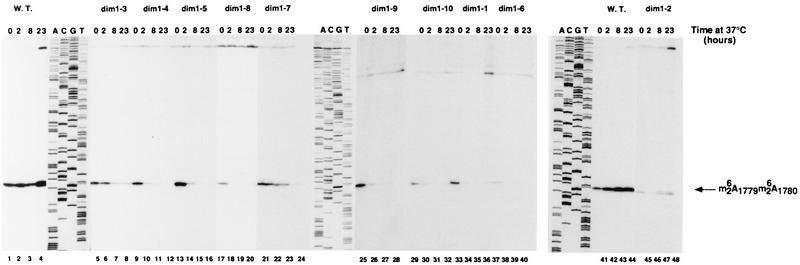
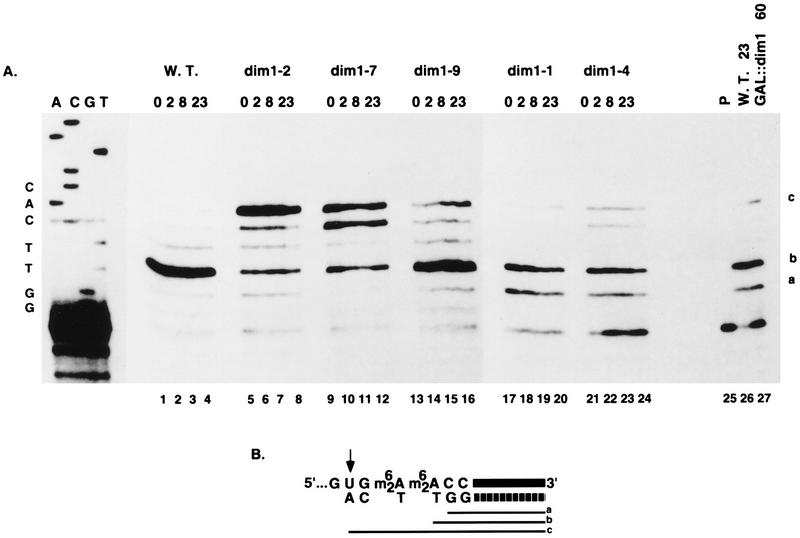
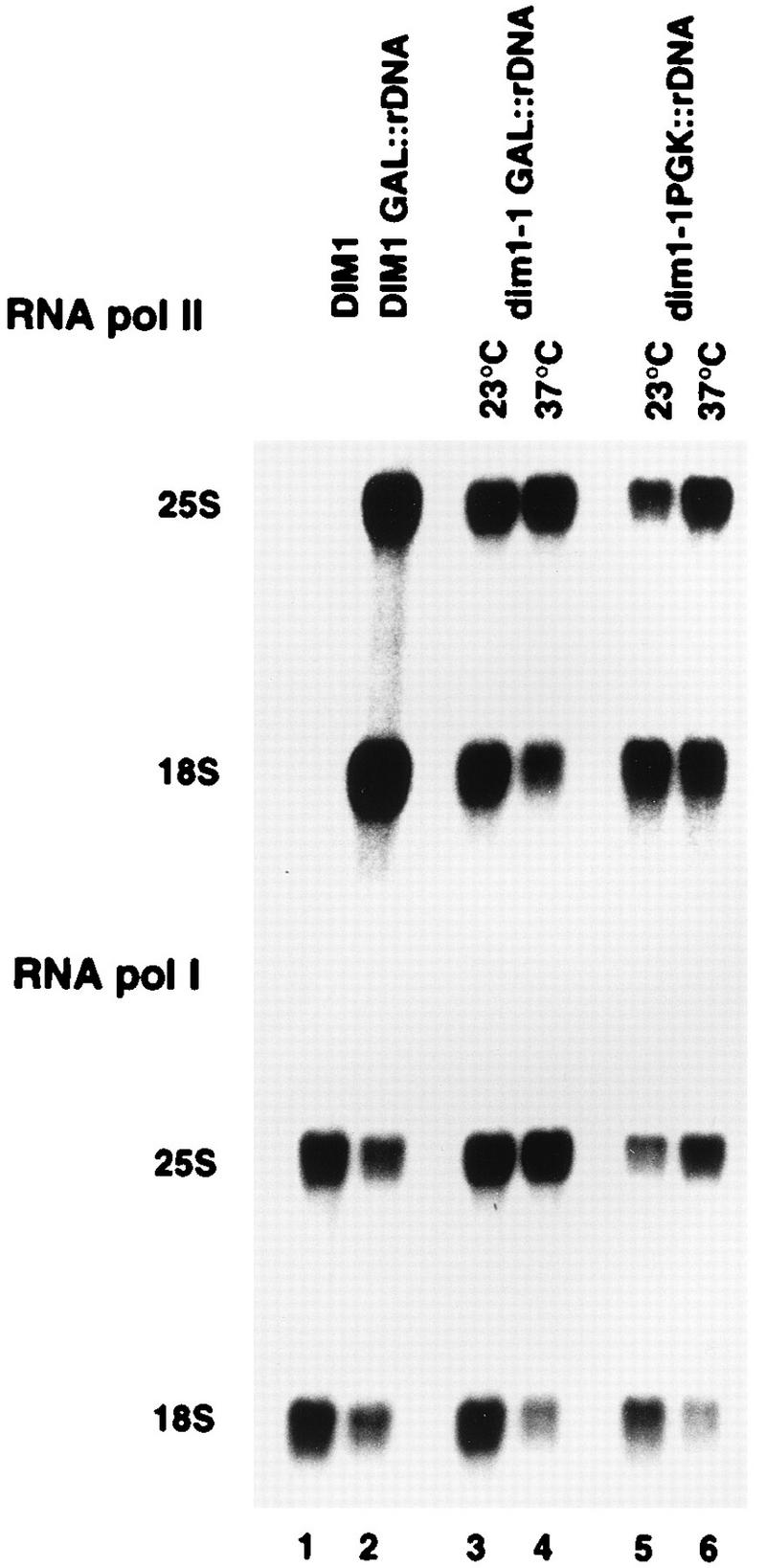
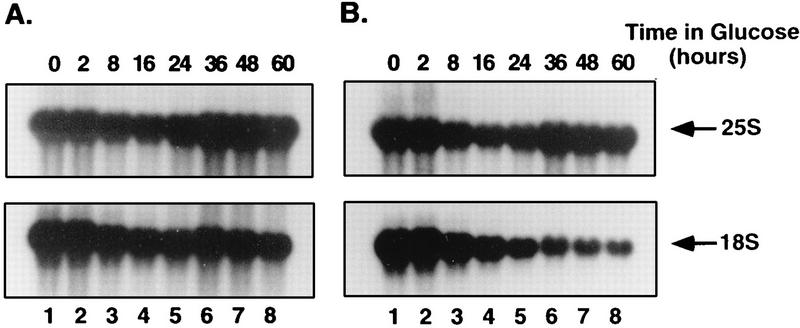
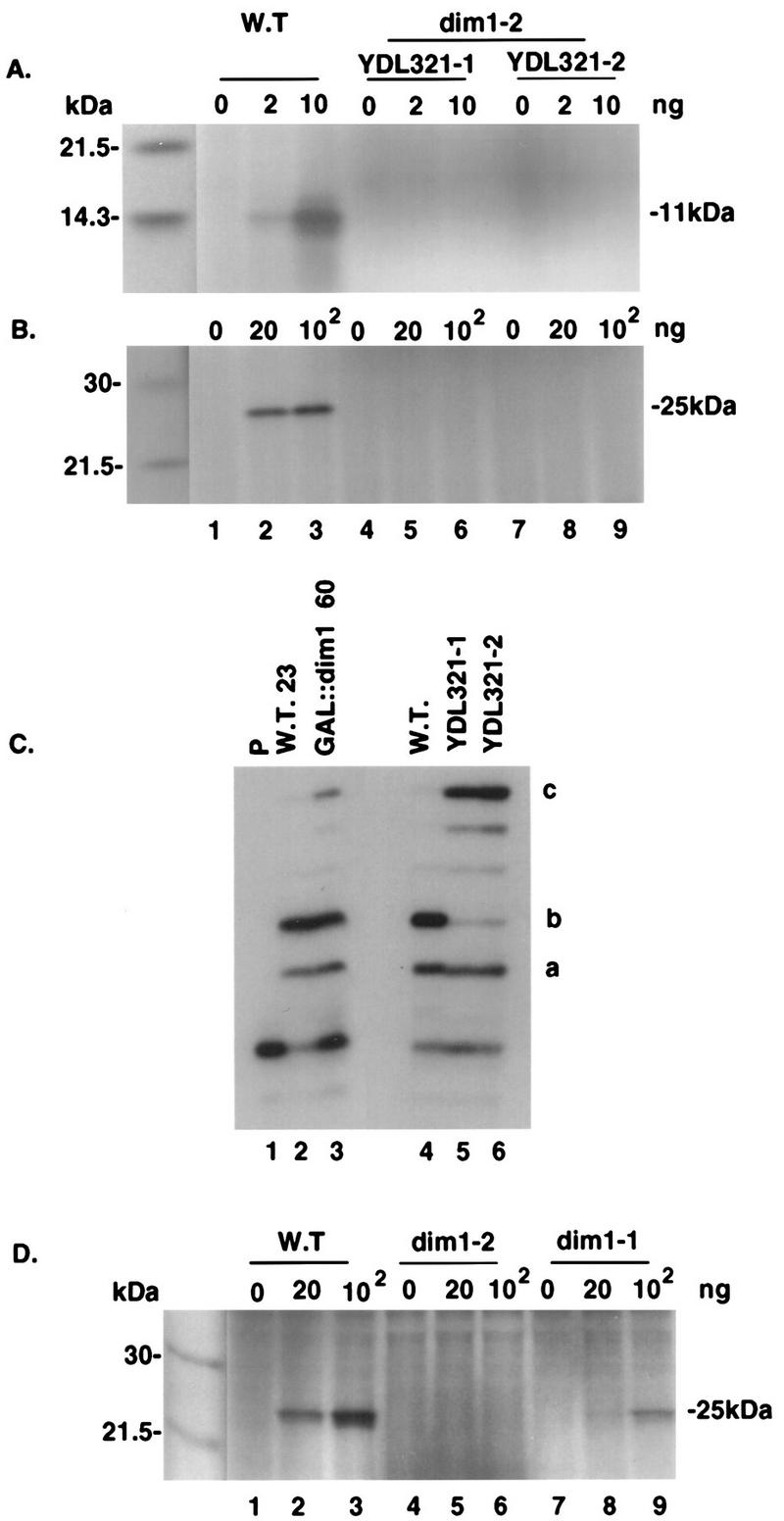
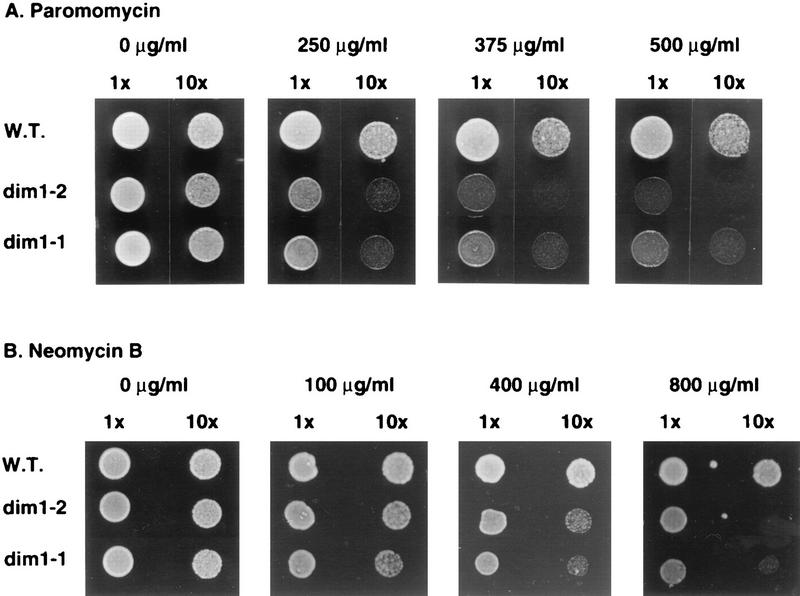
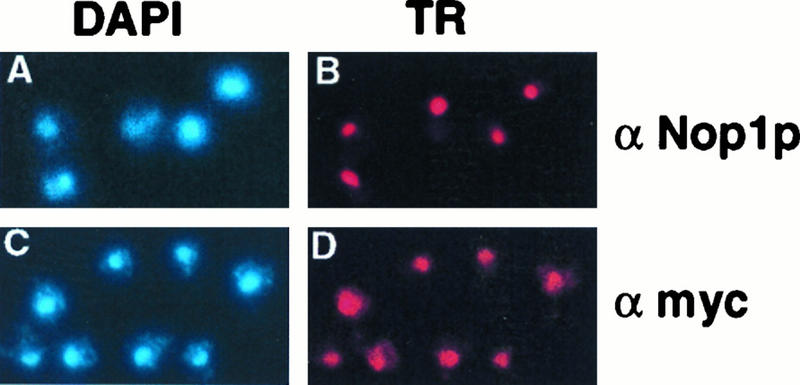
One of the few rRNA modifications conserved between bacteria and eukaryotes is the base dimethylation present at the 3' end of the small subunit rRNA. In the yeast Saccharomyces cerevisiae, this modification is carried out by Dim1p. We previously reported that genetic depletion of Dim1p not only blocked this modification but also strongly inhibited the pre-rRNA processing steps that lead to the synthesis of 18S rRNA. This prevented the formation of mature but unmodified 18S rRNA. The processing steps inhibited were nucleolar, and consistent with this, Dim1p was shown to localize mostly to this cellular compartment. dim1-2 was isolated from a library of conditionally lethal alleles of DIM1. In dim1-2 strains, pre-rRNA processing was not affected at the permissive temperature for growth, but dimethylation was blocked, leading to strong accumulation of nondimethylated 18S rRNA. This demonstrates that the enzymatic function of Dim1p in dimethylation can be separated from its involvement in pre-rRNA processing. The growth rate of dim1-2 strains was not affected, showing the dimethylation to be dispensable in vivo. Extracts of dim1-2 strains, however, were incompetent for translation in vitro. This suggests that dimethylation is required under the suboptimal in vitro conditions but only fine-tunes ribosomal function in vivo. Unexpectedly, when transcription of pre-rRNA was driven by a polymerase II PGK promoter, its processing became insensitive to temperature-sensitive mutations in DIM1 or to depletion of Dim1p. This observation, which demonstrates that Dim1p is not directly required for pre-rRNA processing reactions, is consistent with the inhibition of pre-rRNA processing by an active repression system in the absence of Dim1p.
Figures

References
-
- Berben G, Dumont J, Gilliquet V, Bolle P-A, Hilger F. The YDp plasmids: a uniform set of vectors bearing versatile gene disruption cassettes for Saccharomyces cerevisiae. Yeast. 1991;7:475–477. - PubMed
-
- Boeke J D, Lacroute F, Fink G R. A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol Gen Genet. 1984;197:345–346. - PubMed
-
- Bonneaud N, Ozier-Kalogeropoulos O, Li G, Labouesse M, Minvielle-Sebastia L, Lacroute F. A family of low and high copy replicative, integrative and single-stranded S. cerevisiae/E. coli shuttle vectors. Yeast. 1991;7:609–615. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases