

Assembly of lampbrush chromosomes from sperm chromatin
- PMID: 9529374
- PMCID: PMC25301
- DOI: 10.1091/mbc.9.4.733
Assembly of lampbrush chromosomes from sperm chromatin
Abstract
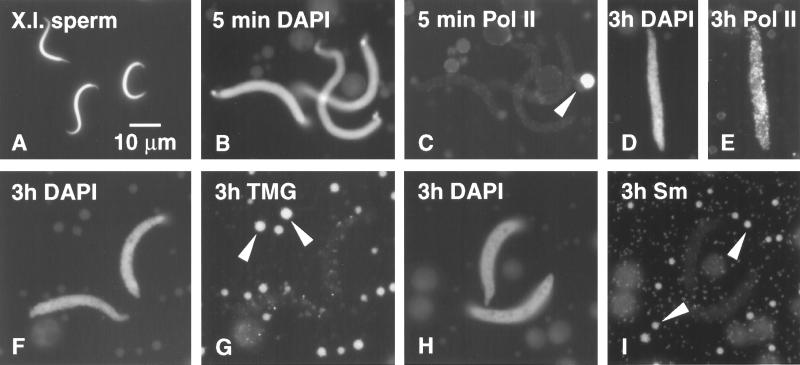
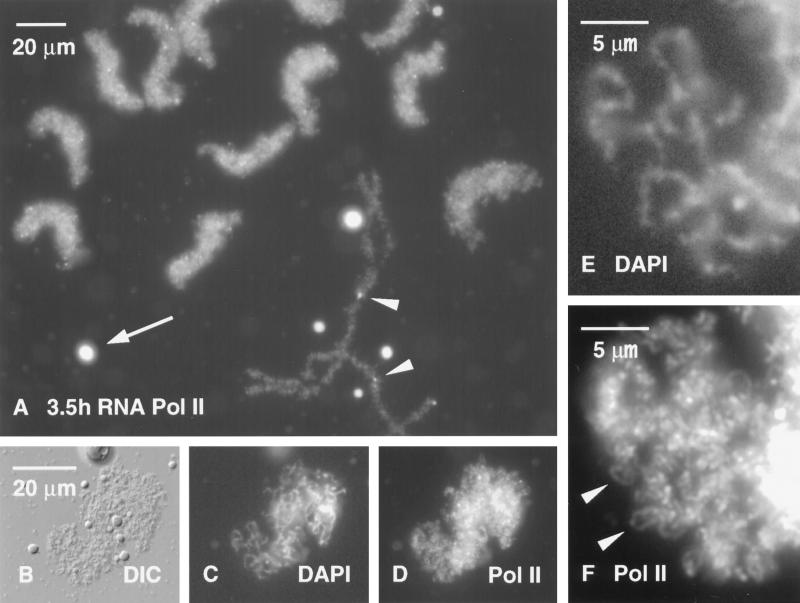
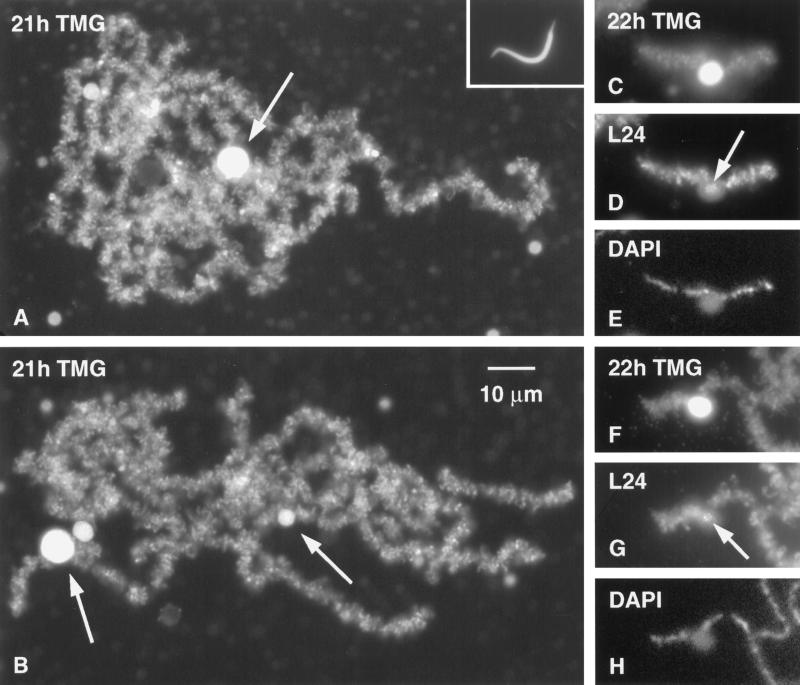
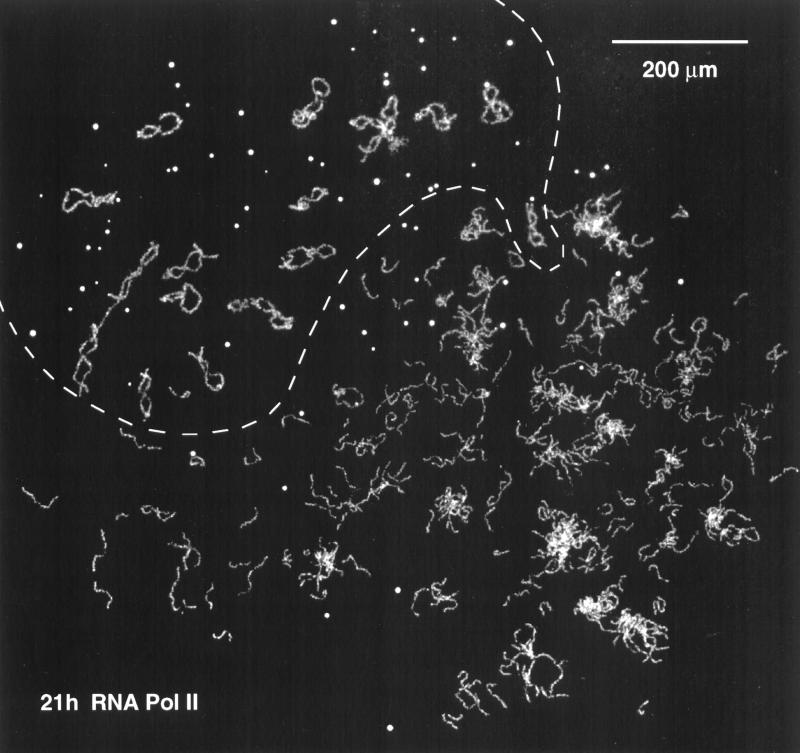
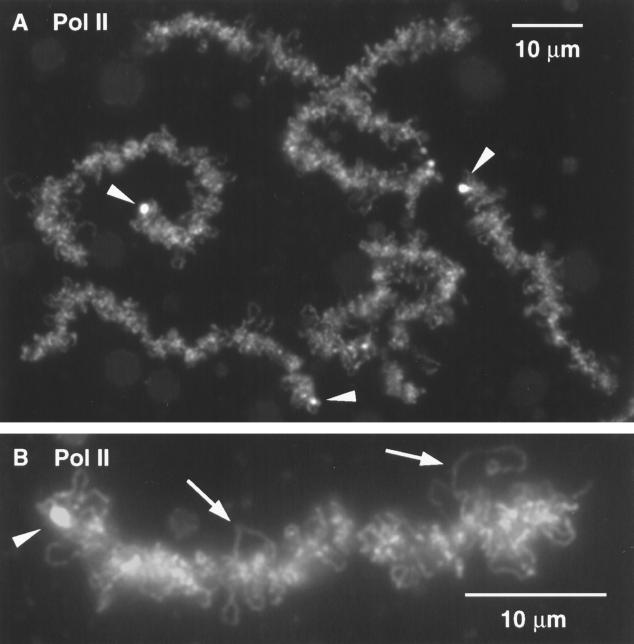
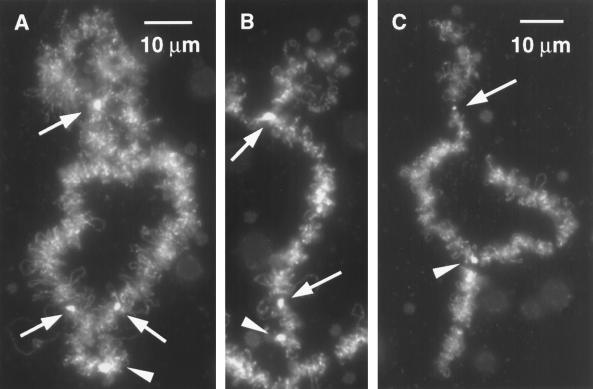
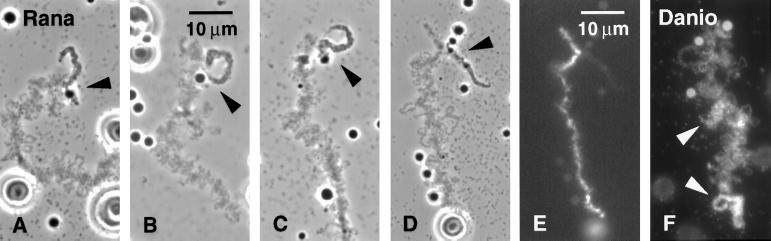
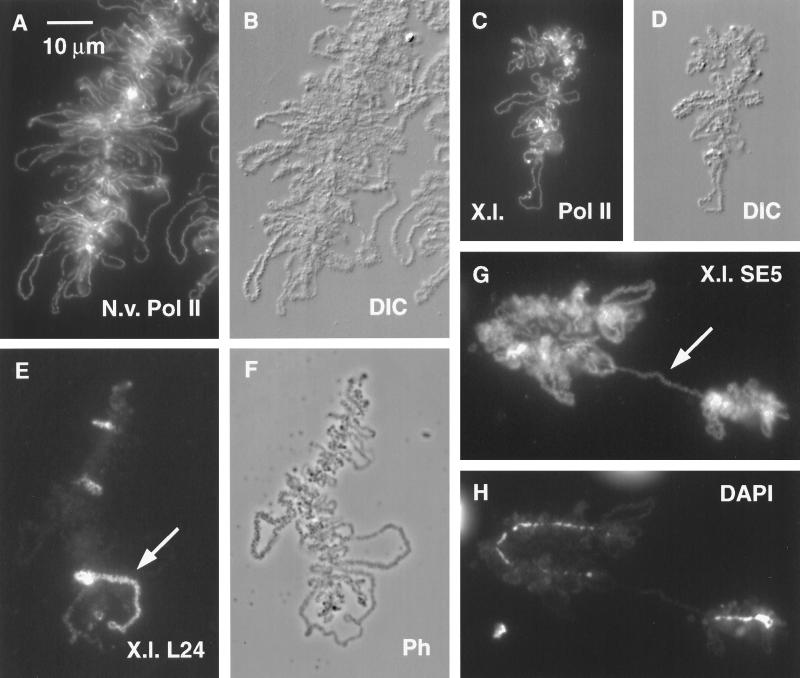
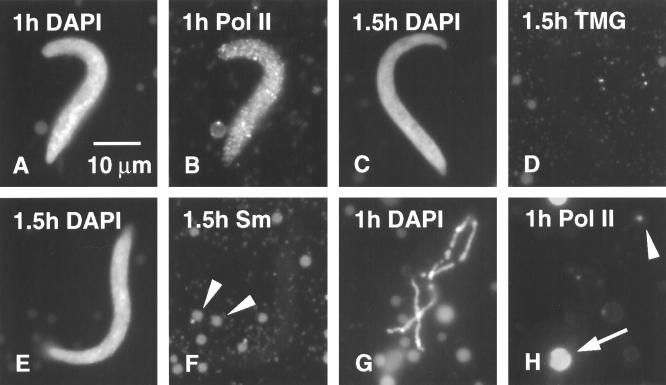
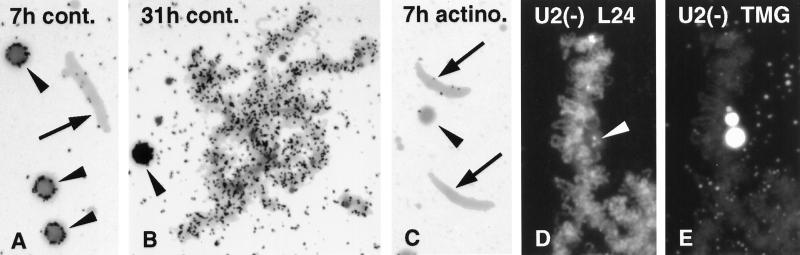
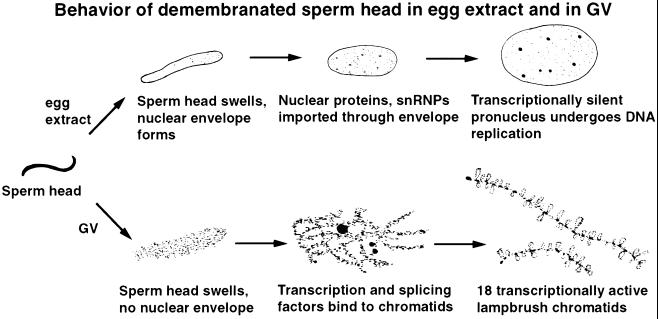
We have examined the behavior of demembranated sperm heads when injected into the germinal vesicle (GV) of amphibian oocytes. Xenopus sperm heads injected into Xenopus GVs swelled immediately and within hours began to stain with an antibody against RNA polymerase II (Pol II). Over time each sperm head became a loose mass of chromosome-like threads, which by 24-48 h resolved into individually recognizable lampbrush chromosomes (LBCs). Although LBCs derived from sperm are unreplicated single chromatids, their morphology and immunofluorescent staining properties were strikingly similar to those of the endogenous lampbrush bivalents. They displayed typical transcriptionally active loops extending from an axis of condensed chromomeres, as well as locus-specific "landmarks. " Experiments with [3H]GTP and actinomycin D demonstrated that transcription was not necessary for the initial swelling of the sperm heads and acquisition of Pol II but was required for maintenance of the lampbrush loops. Splicing was not required at any stage during formation of sperm LBCs. When Xenopus sperm heads were injected into GVs of the newt Notophthalmus, the resulting sperm LBCs displayed very long loops with pronounced Pol II axes, like those of the endogenous newt LBCs; as expected, they stained with antibodies against newt-specific proteins. Other heterologous injections, including sperm heads of the frog Rana pipiens and the zebrafish Danio rerio in Xenopus GVs, confirm that LBCs can be derived from taxonomically distant organisms. The GV system should help identify both cis- and trans-acting factors needed to convert condensed chromatin into transcriptionally active LBCs. It may also be useful in producing cytologically analyzable chromosomes from organisms whose oocytes do not go through a typical lampbrush phase or cannot be manipulated by current techniques.
Figures
References
-
- Almouzni G, Wolffe AP. Nuclear assembly, structure, and function: the use of Xenopus in vitro systems. Exp Cell Res. 1993;205:1–15. - PubMed
-
- Birkenmeier EH, Brown DD, Jordan E. A nuclear extract of Xenopus laevis oocytes that accurately transcribes 5S RNA genes. Cell. 1978;15:1077–1086. - PubMed
-
- Bona M, Scheer U, Bautz EKF. Antibodies to RNA polymerase II (B) inhibit transcription in lampbrush chromosomes after microinjection into living amphibian oocytes. J Mol Biol. 1981;151:81–89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
