

New alleles of the yeast MPS1 gene reveal multiple requirements in spindle pole body duplication
- PMID: 9529376
- PMCID: PMC25304
- DOI: 10.1091/mbc.9.4.759
New alleles of the yeast MPS1 gene reveal multiple requirements in spindle pole body duplication
Abstract
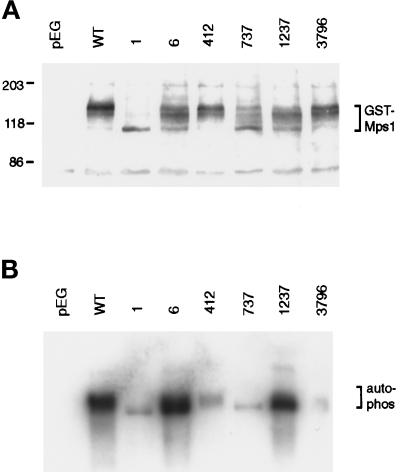
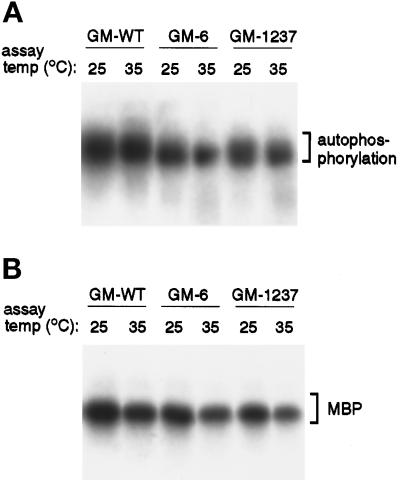
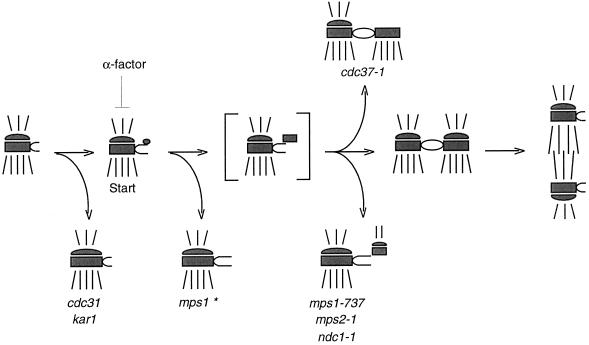
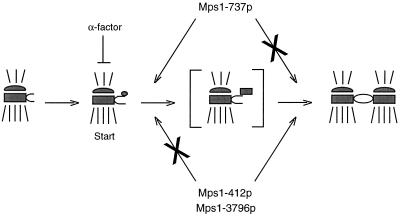
In Saccharomyces cerevisiae, the Mps1p protein kinase is critical for both spindle pole body (SPB) duplication and the mitotic spindle assembly checkpoint. The mps1-1 mutation causes failure early in SPB duplication, and because the spindle assembly checkpoint is also compromised, mps1-1 cells proceed with a monopolar mitosis and rapidly lose viability. Here we report the genetic and molecular characterization of mps1-1 and five new temperature-sensitive alleles of MPS1. Each of the six alleles contains a single point mutation in the region of the gene encoding the protein kinase domain. The mutations affect several residues conserved among protein kinases, most notably the invariant glutamate in subdomain III. In vivo and in vitro kinase activity of the six epitope-tagged mutant proteins varies widely. Only two display appreciable in vitro activity, and interestingly, this activity is not thermolabile under the assay conditions used. While five of the six alleles cause SPB duplication to fail early, yielding cells with a single SPB, mps1-737 cells proceed into SPB duplication and assemble a second SPB that is structurally defective. This phenotype, together with the observation of intragenic complementation between this unique allele and two others, suggests that Mps1p is required for multiple events in SPB duplication.
Figures



References
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. New York: John Wiley & Sons; 1994.
-
- Byers B, Goetsch L. Duplication of spindle plaques and integration of the yeast cell cycle. Cold Spring Harbor Symp Quant Biol. 1974;38:123–131. - PubMed
-
- Carr AM, MacNeill SA, Hayles J, Nurse P. Molecular cloning and sequence analysis of mutant alleles of the fission yeast cdc2+ protein kinase gene: implications for cdc2 protein structure and function. Mol Gen Genetics. 1989;218:41–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
