

Homology-dependent gene silencing in Paramecium
- PMID: 9529389
- PMCID: PMC25319
- DOI: 10.1091/mbc.9.4.931
Homology-dependent gene silencing in Paramecium
Abstract
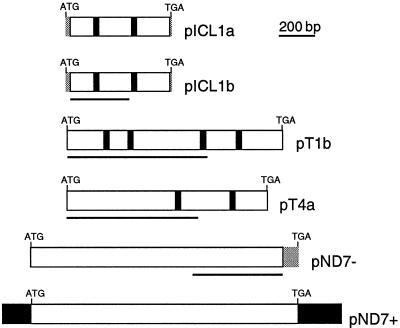
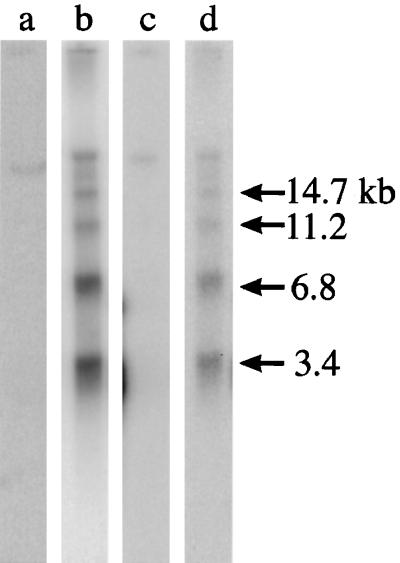
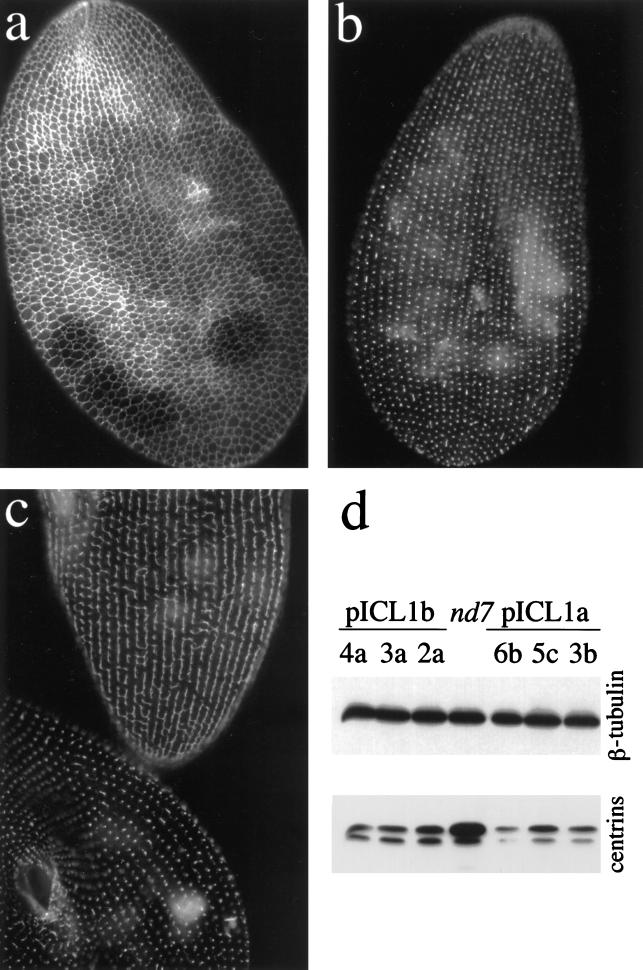
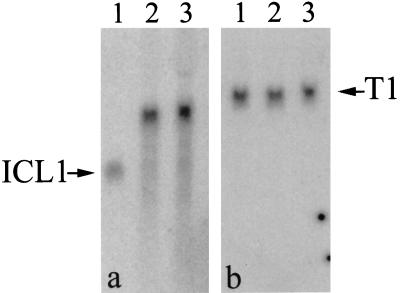
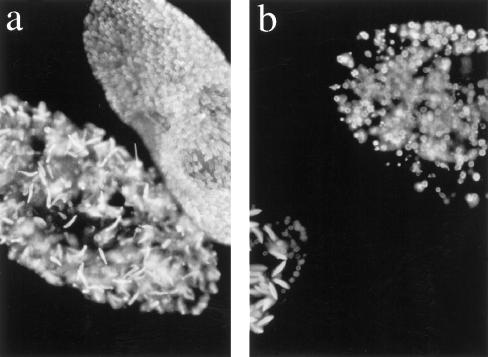
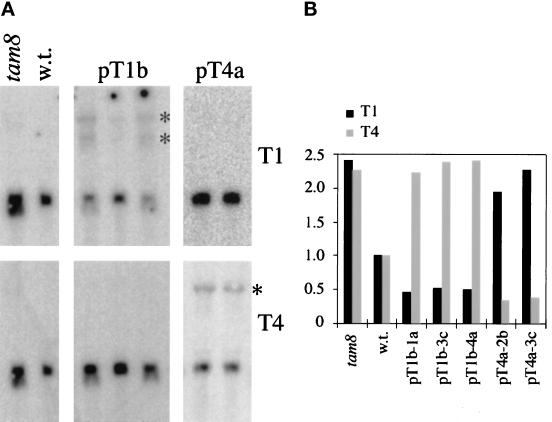
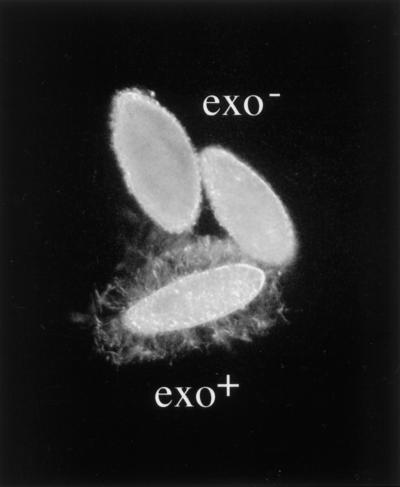
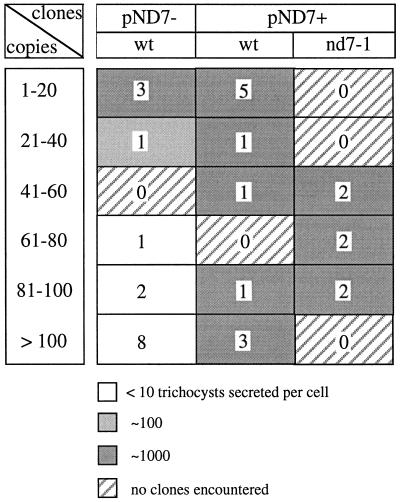
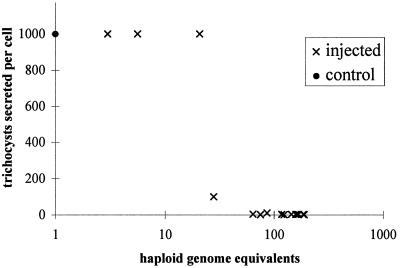
Microinjection at high copy number of plasmids containing only the coding region of a gene into the Paramecium somatic macronucleus led to a marked reduction in the expression of the corresponding endogenous gene(s). The silencing effect, which is stably maintained throughout vegetative growth, has been observed for all Paramecium genes examined so far: a single-copy gene (ND7), as well as members of multigene families (centrin genes and trichocyst matrix protein genes) in which all closely related paralogous genes appeared to be affected. This phenomenon may be related to posttranscriptional gene silencing in transgenic plants and quelling in Neurospora and allows the efficient creation of specific mutant phenotypes thus providing a potentially powerful tool to study gene function in Paramecium. For the two multigene families that encode proteins that coassemble to build up complex subcellular structures the analysis presented herein provides the first experimental evidence that the members of these gene families are not functionally redundant.
Figures
References
-
- Adoutte A, Garreau de Loubresse N, Beisson J. Proteolytic cleavage and maturation of the crystalline secretion products of Paramecium. J Mol Biol. 1984;180:1065–1080. - PubMed
-
- Adoutte A. Exocytosis: biogenesis, transport and secretion of trichocysts. In: Görz H-D, editor. Paramecium. Heidelberg: Springer-Verlag; 1988. pp. 325–362.
-
- Baulcombe DC. RNA as a target and an initiator of post-transcriptional gene silencing in transgenic plants. Plant Mol Biol. 1996;32:79–88. - PubMed
-
- Beisson J, Rossignol M. Molecular Biology of Nucleocytoplasmic Relationships. The Netherlands: Elsevier; 1975. Movements and positioning of organelles in Paramecium aurelia; pp. 291–294.
-
- Bingham PM. Cosuppression comes to animals. Cell. 1997;90:385–387. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
