

Swi5 controls a novel wave of cyclin synthesis in late mitosis
- PMID: 9529390
- PMCID: PMC25320
- DOI: 10.1091/mbc.9.4.945
Swi5 controls a novel wave of cyclin synthesis in late mitosis
Abstract
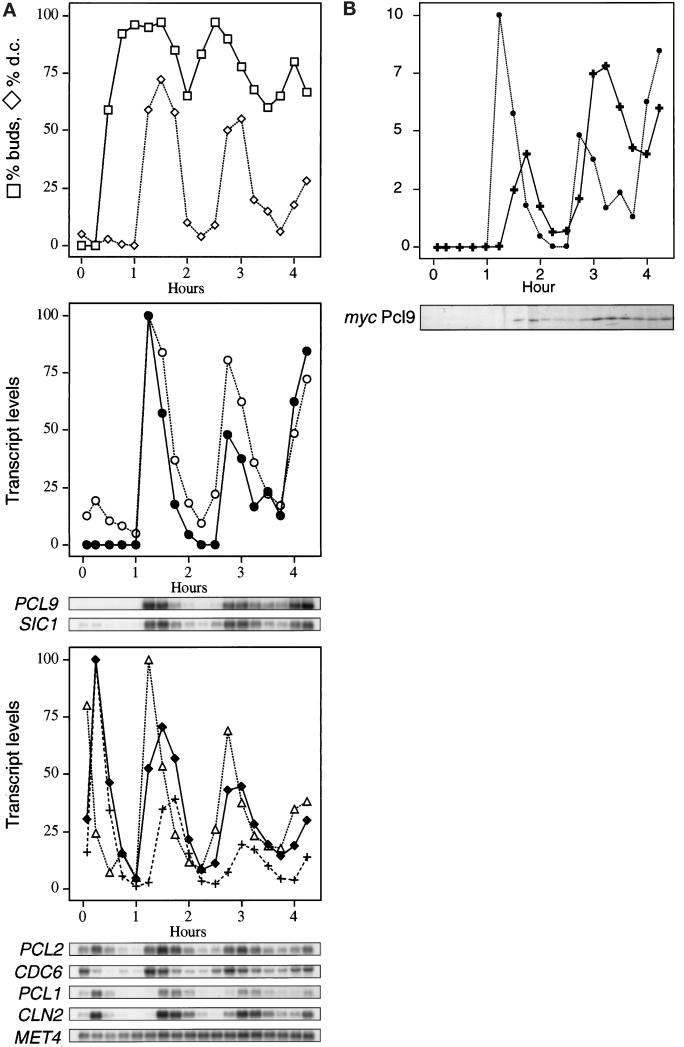
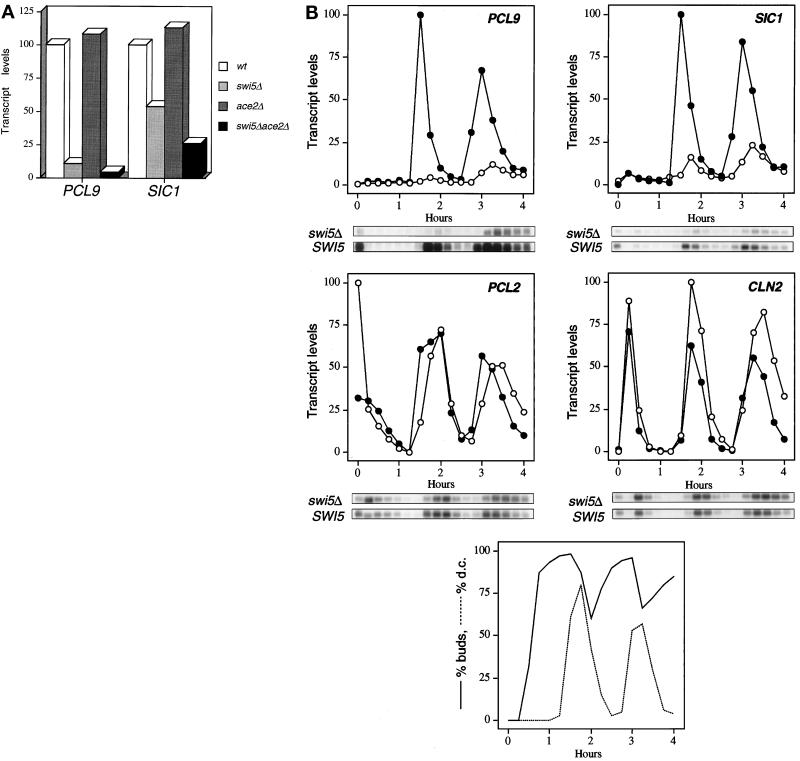
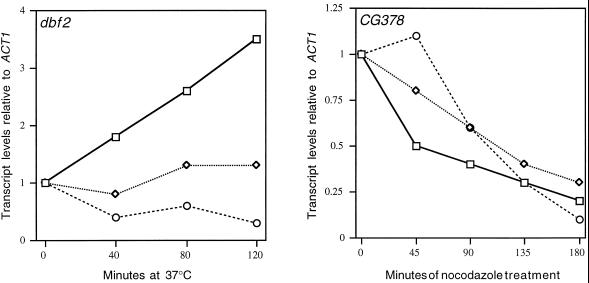
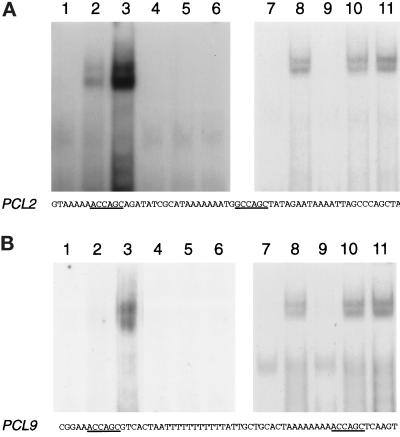
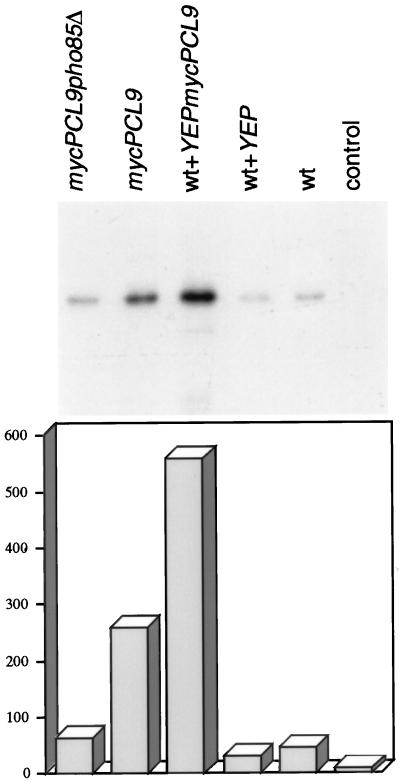
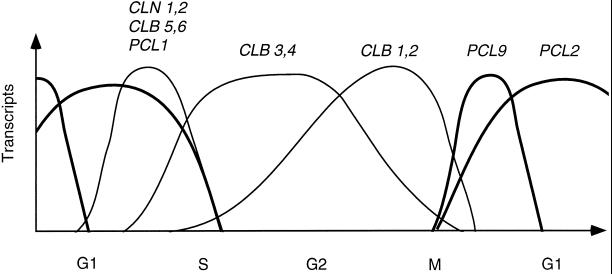
We have shown previously that the Swi5 transcription factor regulates the expression of the SIC1 Cdk inhibitor in late mitosis. This suggests that Swi5 might control other genes with roles in ending mitosis. We identified a gene with a Swi5-binding site in the promoter that encoded a protein with high homology to Pcl2, a cyclin-like protein that associates with the Cdk Pho85. This gene, PCL9, is indeed regulated by Swi5 in late M phase, the only cyclin known to be expressed at this point in the cell cycle. The Pcl9 protein is associated with a Pho85-dependent protein kinase activity, and the protein is unstable with peak levels occurring in late M phase. PCL2 is already known to be expressed in late G1 and we find that, in addition, it is also regulated by Swi5 in telophase. The expression of PCL2 and PCL9 at this stage of the cell cycle implies a role for the Pho85 Cdk at the end of mitosis. Consistent with this a synthetic interaction was observed between pho85delta and strains deleted for SIC1, SWI5, and SPO12. These and other studies support the notion that the M/G1 switch is a major cell cycle transition.
Figures

References
-
- Bobola N, Jansen R-P, Shin TH, Nasmyth K. Asymmetric accumulation of Ash1p in post anaphase nuclei depends on a myosin and restricts yeast mating-type switching to mother cells. Cell. 1996;84:699–709. - PubMed
-
- Cocker JH, Piatti S, Santocanale C, Nasmyth K, Diffley JF. An essential role for the Cdc6 protein in forming the pre-replicative complexes of budding yeast. Nature. 1996;379:180–182. - PubMed
-
- Cohen-Fix O, Peters JM, Kirschner MW, Koshland D. Anaphase initiation in Saccharomyces cerevisiae is controlled by the APC-dependent degradation of the anaphase inhibitor Pds1p. Genes Dev. 1996;10:3081–3093. - PubMed
-
- Dohrmann PR, Butler G, Tamai K, Dorland S, Greene JR. Parallel pathways of gene regulation: homologous regulators SWI5 and ACE2 differentially control transcription of HO and chitinase. Genes Dev. 1992;6:93–104. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
