

Three drought-responsive members of the nonspecific lipid-transfer protein gene family in Lycopersicon pennellii show different developmental patterns of expression
- PMID: 9536064
- PMCID: PMC35054
- DOI: 10.1104/pp.116.4.1461
Three drought-responsive members of the nonspecific lipid-transfer protein gene family in Lycopersicon pennellii show different developmental patterns of expression
Abstract
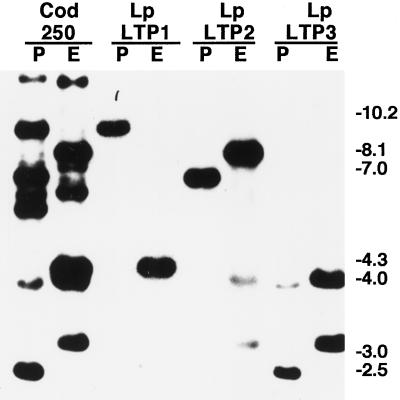
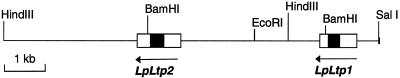
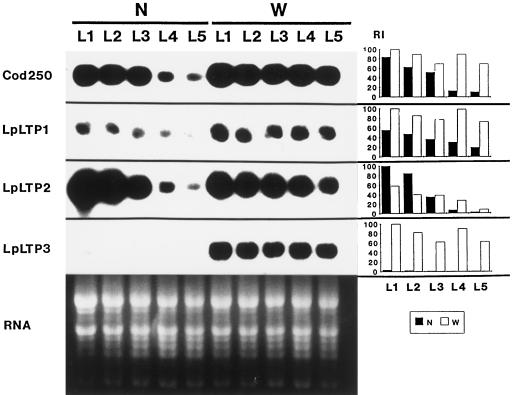
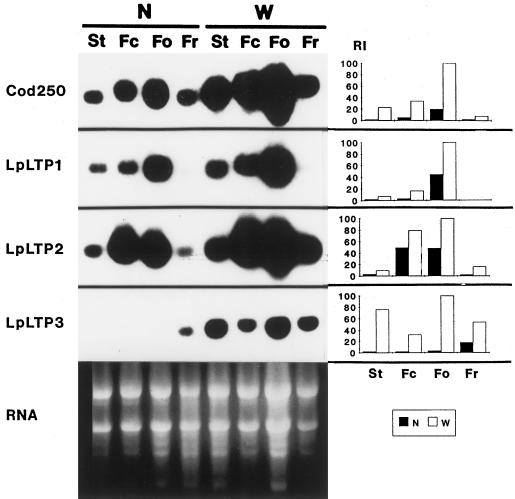
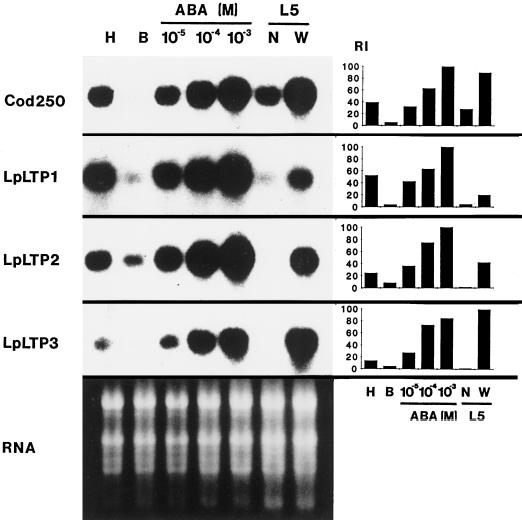
Genomic clones of two nonspecific lipid-transfer protein genes from a drought-tolerant wild species of tomato (Lycopersicon pennellii Corr.) were isolated using as a probe a drought- and abscisic acid (ABA)-induced cDNA clone (pLE16) from cultivated tomato (Lycopersicon esculentum Mill.). Both genes (LpLtp1 and LpLtp2) were sequenced and their corresponding mRNAs were characterized; they are both interrupted by a single intron at identical positions and predict basic proteins of 114 amino acid residues. Genomic Southern data indicated that these genes are members of a small gene family in Lycopersicon spp. The 3'-untranslated regions from LpLtp1 and LpLtp2, as well as a polymerase chain reaction-amplified 3'-untranslated region from pLE16 (cross-hybridizing to a third gene in L. pennellii, namely LpLtp3), were used as gene-specific probes to describe expression in L. pennellii through northern-blot analyses. All LpLtp genes were exclusively expressed in the aerial tissues of the plant and all were drought and ABA inducible. Each gene had a different pattern of expression in fruit, and LpLtp1 and LpLtp2, unlike LpLtp3, were both primarily developmentally regulated in leaf tissue. Putative ABA-responsive elements were found in the proximal promoter regions of LpLtp1 and LpLtp2.
Figures

References
-
- Altschul SF, Warren G, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Arondel V, Tchang F, Baillet B, Vignols F, Grellet F, Delsney M, Kader JC, Puigdomenech P. Multiple mRNA coding for phospholipid-transfer protein from Zea mays arise from alternative splicing. Gene. 1991;99:133–136. - PubMed
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current Protocols in Molecular Biology. New York: John Wiley & Sons; 1996.
-
- Bourgis F, Kader J-C. Lipid-transfer proteins: tools for manipulating membrane lipids. Physiol Plant. 1997;100:78–84.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
