

Accumulation of a clock-regulated transcript during flower-inductive darkness in pharbitis nil
- PMID: 9536066
- PMCID: PMC35056
- DOI: 10.1104/pp.116.4.1479
Accumulation of a clock-regulated transcript during flower-inductive darkness in pharbitis nil
Abstract
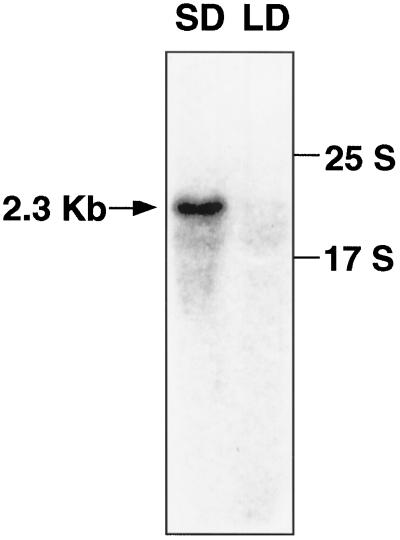
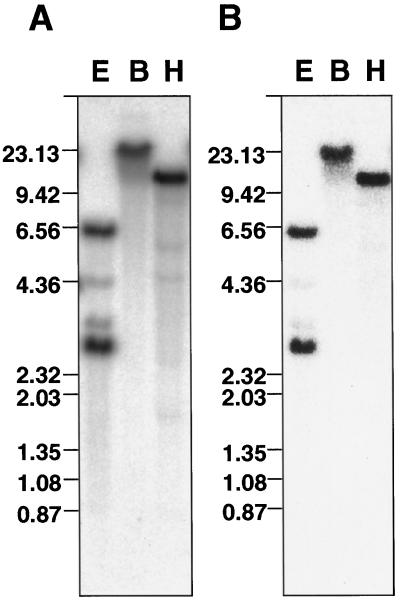
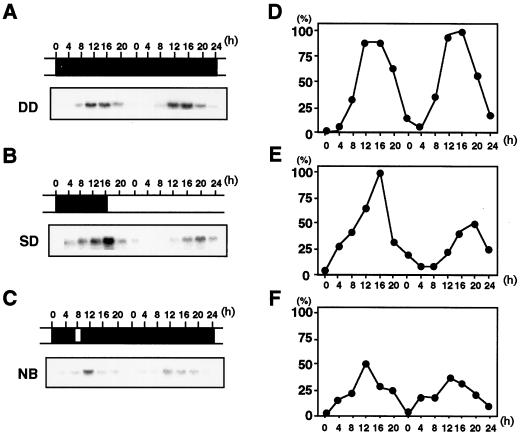
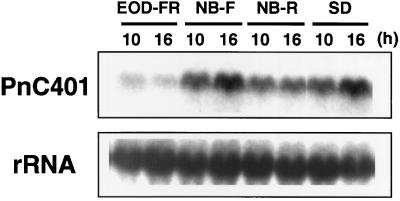
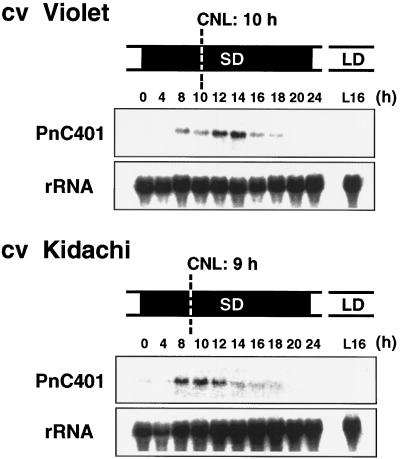
To clarify the molecular basis of the photoperiodic induction of flowering in the short-day plant Pharbitis nil cv Violet, we examined changes in the level of mRNA in cotyledons during the flower-inductive photoperiod using the technique of differential display by the polymerase chain reaction. A transcript that accumulated during the inductive dark period was identified and a cDNA corresponding to the transcript, designated PnC401 (P. nil C401), was isolated. RNA-blot hybridization verified that levels of PnC401 mRNA fluctuated with a circadian rhythm, with maxima between 12 and 16 h after the beginning of the dark period) and minima of approximately 0. This oscillation continued even during an extended dark period but was damped under continuous light. Accumulation of PnC401 mRNA was reduced by a brief exposure to red light at the 8th h of the dark period (night-break treatment) or by exposure to far-red light at the end of the light period (end-of-day far-red treatment). These results suggest that fluctuations in levels of PnC401 mRNA are regulated by phytochrome(s) and a circadian clock and that they are associated with photoperiodic events that include induction of flowering.
Figures

References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Beator J, Kloppstech K. Significance of circadian gene expression in higher plants. Chronobiol Int. 1996;13:319–339. - PubMed
-
- Bernier G. The control of floral evocation and morphogenesis. Annu Rev Plant Physiol Plant Mol Biol. 1988;39:175–219.
-
- Evans LT. Flower induction and the florigen concept. Annu Rev Plant Physiol. 1971;22:365–394.
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
