

The CIRCE element and its putative repressor control cell cycle expression of the Caulobacter crescentus groESL operon
- PMID: 9537357
- PMCID: PMC107072
- DOI: 10.1128/JB.180.7.1632-1641.1998
The CIRCE element and its putative repressor control cell cycle expression of the Caulobacter crescentus groESL operon
Abstract
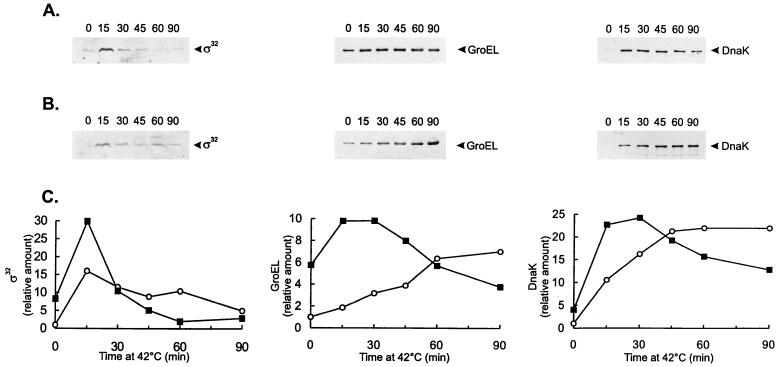
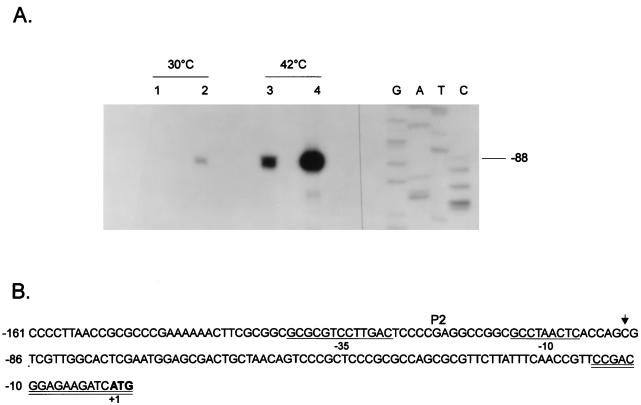
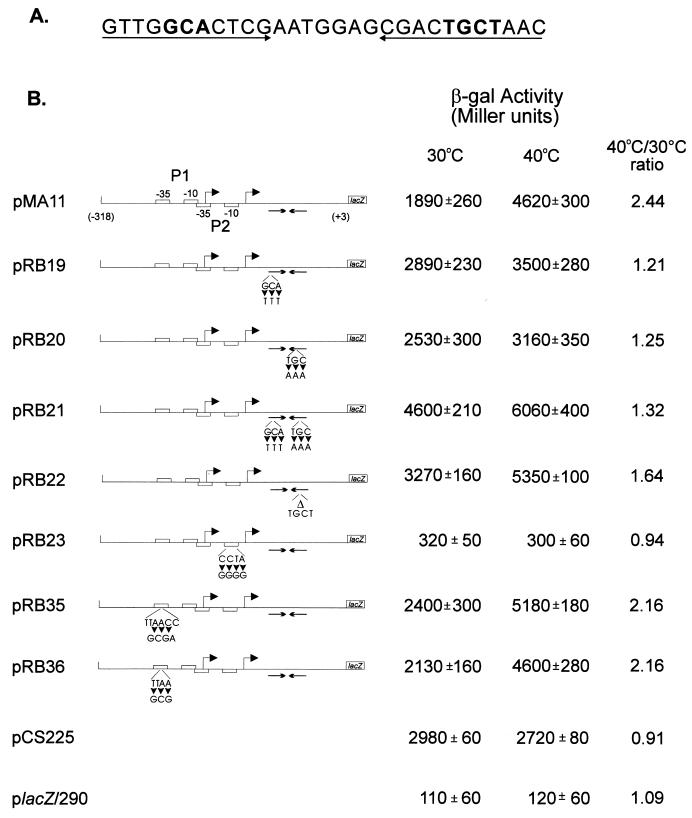
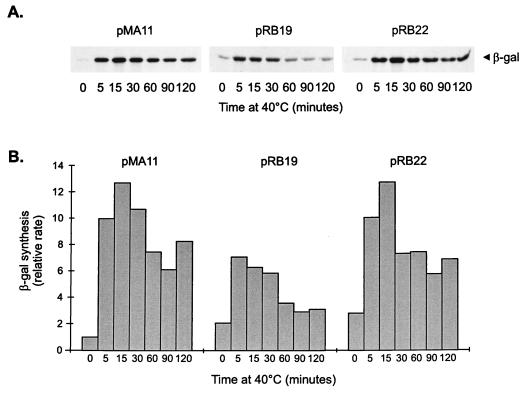
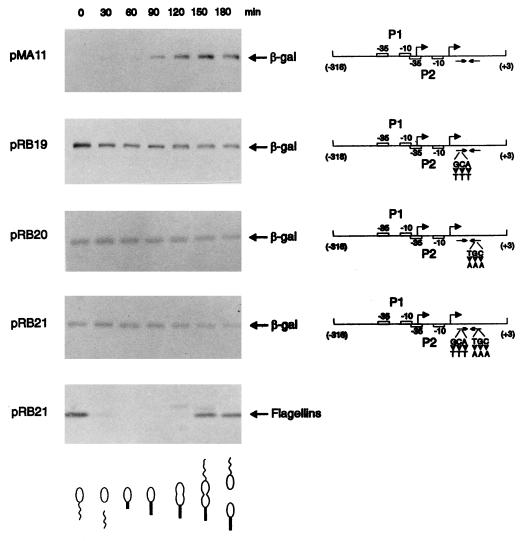
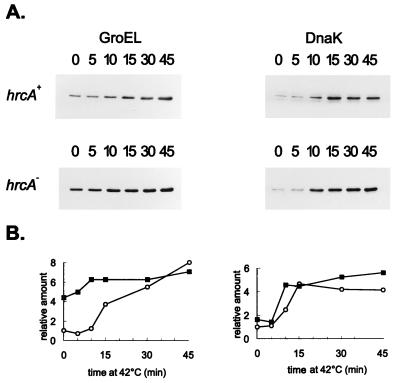
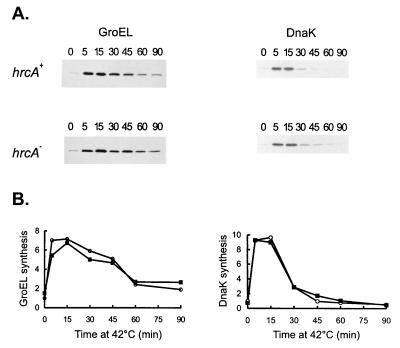
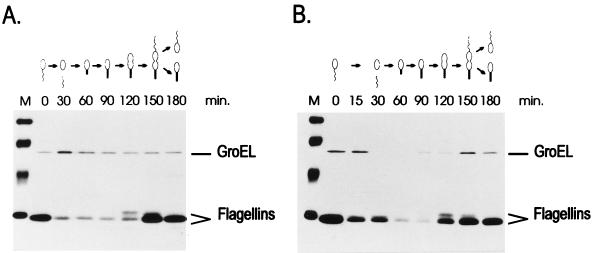
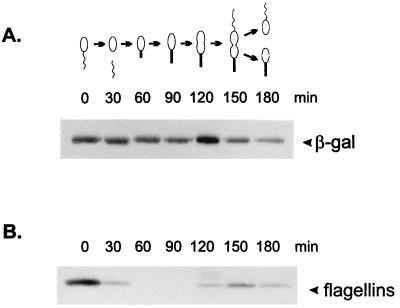
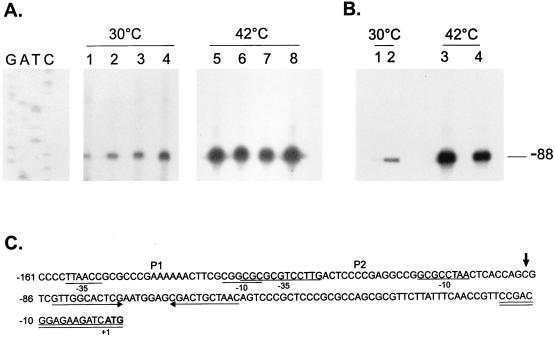
The groESL operon is under complex regulation in Caulobacter crescentus. In addition to strong induction after exposure to heat shock, under physiological growth conditions, its expression is subject to cell cycle control. Transcription and translation of the groE genes occur primarily in predivisional cells, with very low levels of expression in stalked cells. The regulatory region of groESL contains both a sigma32-like promoter and a CIRCE element. Overexpression of C. crescentus sigma32 gives rise to higher levels of GroEL and increased levels of the groESL transcript coming from the sigma32-like promoter. Site-directed mutagenesis in CIRCE has indicated a negative role for this cis-acting element in the expression of groESL only at normal growth temperatures, with a minor effect on heat shock induction. Furthermore, groESL-lacZ transcription fusions carrying mutations in CIRCE are no longer cell cycle regulated. Analysis of an hrcA null strain, carrying a disruption in the gene encoding the putative repressor that binds to the CIRCE element, shows constitutive synthesis of GroEL throughout the Caulobacter cell cycle. These results indicate a negative role for the hrcA gene product and the CIRCE element in the temporal control of the groESL operon.
Figures
References
-
- Antoine R, Locht C. Isolation and molecular characterization of a novel broad-host-range plasmid from Bordetella bronchiseptica with sequence similarities to plasmids from gram-positive organisms. Mol Microbiol. 1992;6:1785–1799. - PubMed
-
- Avedissian M, Gomes S L. Expression of the groESL operon is cell cycle controlled in Caulobacter crescentus. Mol Microbiol. 1996;19:79–89. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
