

doi: 10.1073/pnas.95.8.4264.
The TOR (target of rapamycin) signal transduction pathway regulates the stability of translation initiation factor eIF4G in the yeast Saccharomyces cerevisiae
Affiliations
- PMID: 9539725
- PMCID: PMC22477
- DOI: 10.1073/pnas.95.8.4264
Item in Clipboard
The TOR (target of rapamycin) signal transduction pathway regulates the stability of translation initiation factor eIF4G in the yeast Saccharomyces cerevisiae
Proc Natl Acad Sci U S A.
.
Abstract
Initiation factor eIF4G is an essential protein required for initiation of mRNA translation via the 5' cap-dependent pathway. It interacts with eIF4E (the mRNA 5' cap-binding protein) and serves as an anchor for the assembly of further initiation factors. With treatment of Saccharomyces cerevisiae with rapamycin or with entry of cells into the diauxic phase, eIF4G is rapidly degraded, whereas initiation factors eIF4E and eIF4A remain stable. We propose that nutritional deprivation or interruption of the TOR signal transduction pathway induces eIF4G degradation.
Figures
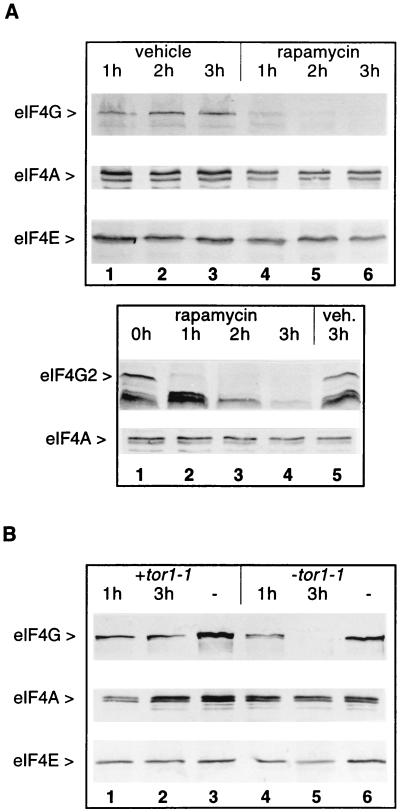
Rapamycin causes the disappearance of eIF4G. (A) Exponentially growing cells from strains CWO4 (Upper) or CBY1.1 (Lower) were treated with rapamycin or vehicle. Aliquots (20 ml) were harvested at the times indicated after addition of the drug (or vehicle). Cell extracts were fractionated by SDS/PAGE, blotted onto nitrocellulose, and decorated with rat polyclonal antibodies against eIF4G (Upper and Lower, first row), eIF4A (Upper and Lower, second row), and eIF4E (Upper, third row). (B) Cell extracts from strain CWO4 transformed with the plasmid pPW2 carrying the dominant tor1-1 allele (lanes 1–3) or a vector without tor1-1 (lanes 4–6) were treated as described in A with rapamycin (lanes 1, 2, 4 and 5) or vehicle (lanes 3 and 6) and analyzed on Western blots for eIF4G (first row), eIF4A (second row), or eIF4E content (third row).
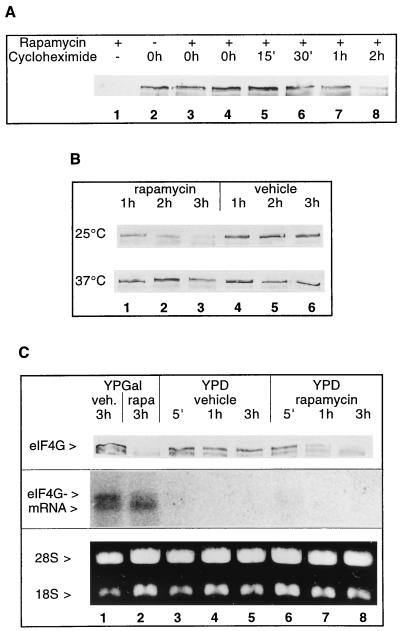
Stability of eIF4G. (A) Effect of rapamycin treatment on eIF4G (strain CWO4) when de novo protein synthesis is blocked with cycloheximide. Rapamycin was added to exponentially growing cells at time point 0 h, except for lane 2 (no rapamycin added) and lane 3 where it was added 15 min after cycloheximide addition. The time points of cycloheximide addition are indicated. The total duration of incubation in the presence of the drugs was 3 h. Shown are Western blots from extracts decorated for eIF4G. (B) Effect of rapamycin (lanes 1–3) or vehicle (lanes 4–6) on eIF4G stability in the mutant strain PDY4 (cdc63-1) grown at 25°C (first row) and after shift from 25°C to 37°C (second row). Time points after addition of the drug (or vehicle) are indicated. Shown are Western blots decorated for eIF4G. (C) Exponentially growing cells in YPGal (strain CBY12) were shifted to fresh medium containing YPGal (lanes 1 and 2) or YPD (lanes 3–8) and incubated with rapamycin or vehicle for the times indicated. First row shows Western blot decorated for eIF4G. Equal volumes of cell cultures were loaded. Second row shows Northern blot of total RNA isolated from cells harvested at the time points indicated and probed with a cDNA corresponding to the whole TIF4631 ORF (23). Third row shows ethidium bromide staining of 28S and 18S rRNAs used for the Northern blot analysis.
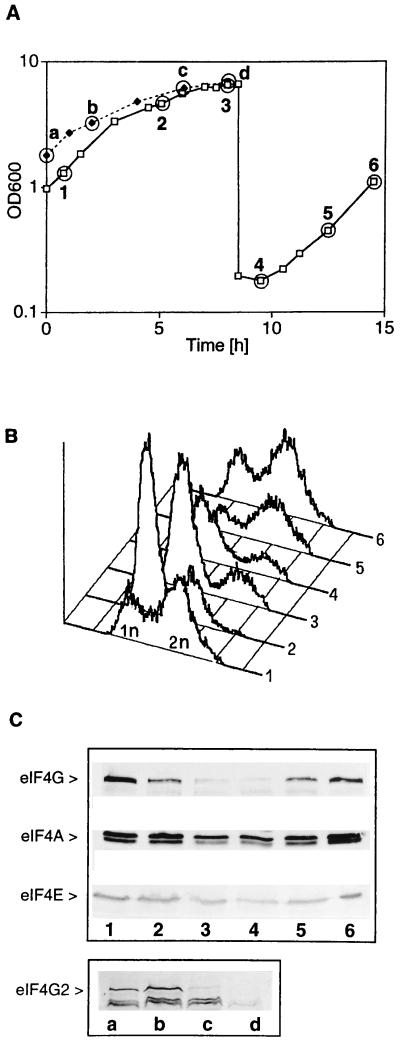
eIF4G degradation is induced upon entry into the diauxic phase. (A) Strain CWO4 was allowed to grow to the diauxic phase (points 2 and 3), diluted into fresh medium, and allowed to resume exponential growth (points 4–6). Strain CBY1.1 (TIF4631 deletion) was allowed to grow to the diauxic phase (points c and d). Time points indicated (–6) correspond to CWO4 cells harvested at a cell density (OD600) of 1.302 (time point 1), 4.66 (time point 2), 6.6 (time point 3), 0.178 (time point 4), 0.445 (time point 5), and 1.10 (time point 6) and to CBY1.1 cells with an OD600 of 1.81 (time point a), 3.29 (time point b), 6.19 (time point c), and 6.88 (time point d). (B) Flow cytometry analysis (graphs) of CWO4 cells harvested at time points 1–6 in A. (C) Western blots of extracts from CWO4 cells harvested at time points 1–6 (lanes 1–6) and CBY1.1 (lanes a–d) and decorated with antibodies against eIF4G, eIF4A, and eIF4E.
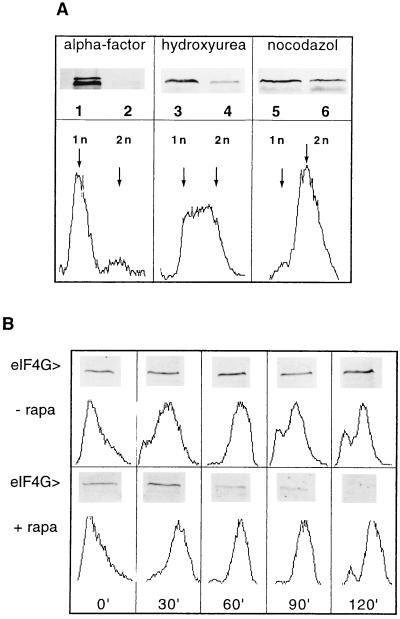
eIF4G degradation is not confined to a specific stage of the cell cycle. (A Upper) Exponentially growing cells (strain ASZ1) were arrested at an OD600 of 0.2–0.3/ml by treating them either with α-factor for 2 h (lanes 1 and 2), with hydroxyurea for 2 h (lanes 3 and 4), or with nocodazole for 1 h (lanes 5 and 6). Vehicle (lanes 1, 3, and 5) or rapamycin (lanes 2, 4, and 6) was added to the arrested cultures and incubation was continued for 2 h (except nocodazole-treated cells for 1.5 h). Cell extracts were analyzed on Western blots for eIF4G content (lanes 1–6). (A Lower)The graphs show the flow cytometry analysis of cells arrested with the different drugs. (B) Exponentially growing ASZ1 cells were arrested with α-factor (2 h treatment) and subsequently released by incubating with fresh YPD in the absence (Upper) or in the presence of rapamycin (Lower). At the time points indicated (0, 30, 60, 90, and 120 min) cell cultures (10 ml) were harvested, aliquots were prepared for flow cytometry analysis, and cell extracts were analyzed for eIF4G content by Western blotting and immunodecoration.
References
-
- Werner-Washburne M, Braun E L, Crawford M E, Peck V M. Mol Microbiol. 1996;19:1159–1166. - PubMed
-
- Pringle J R, Hartwell L H. In: The Saccharomyces Cerevisiae Life Cycle. Strathern J N, Jones E W, Broach J R, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1992. pp. 97–142.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
